

Beljakovine g struktura, vrste in funkcije
- 4595
- 514
- Mr. Shane Larkin
The G tudi Guaninski nukleotidni vezavni proteini, So beljakovine, povezane z plazemsko membrano, ki pripada družini "spojkov" signalov, ki imajo pomembne funkcije v številnih procesih transdukcije signala v evkariontskih organizmih.
V literaturi so G proteini opisani kot binarna molekularna stikala, saj njihova biološka aktivnost določajo spremembe njihove strukture, ki jih daje nukleotidna vrsta, na katero se lahko združijo: gvanozin nukleotidi (difosfat (GDP) in tiposfat (GTP)).
Struktura proteina RAS, monomernega G proteina (vir: Mark 'Absturz' [javna domena] prek Wikimedia Commons) Na splošno jih aktivirajo receptorji iz družine beljakovin, znanih kot GPCR, povezani z receptorji (GPCR, angleščina "G-beljakovinski sprejemniki "), ki prejmejo zunanji začetni signal in naredijo konformacijske spremembe, ki sprožijo aktivacijo, kar pozneje prevede v aktivacijo drugega efektorskega proteina.
Nekateri avtorji menijo, da so se kodirajoči geni za to družino beljakovin razvijali v podvajanje in razhajanje skupnega gena prednikov, katerega izdelek je bil rafiniran in specializiran vedno več.
Med najrazličnejšimi celičnimi funkcijami, ki jih imajo ti proteini.
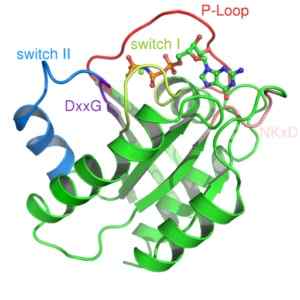
Iz te vrste beljakovin sta bila opisana dva razreda: majhni G proteini in heterotrofimerni G proteini. Prvo tridimenzionalno strukturo G proteina smo dobili več kot desetletje od majhnega G proteina, znanega kot RAS.
[TOC]
Struktura
Strukturno gledano sta prepoznana dve vrsti G -proteinov: majhni G proteini in heterotrimični G proteini, veliko bolj zapleteni.
Majhne G proteine tvorijo en sam polipeptid okoli 200 aminoazijskih odpadkov in približno 20-40 kDa, v njegovi strukturi pa je ohranjena katalitična domena (domena G), sestavljena iz petih α propelerjev, šest β-utripanih listov in pet polipeptida zanke.
Heterotrofimerni G proteini so na drugi strani celovito membransko beljakovine, ki so sestavljeni iz treh polipeptidnih verig, znanih kot α, β in γ podenote.
-Podenota α tehta med 40 in 52 kDa, ima gvanin nukleotidno vezavno območje in ima aktivnost GTPASA za hidrolize med GTP fosfatnimi skupinami.
Α podenote različnih beljakovin G imajo nekatere strukturne domene, kot sta Union GTP in hidroliza, vendar se na mestih vezave receptorjev in efektorskih beljakovin zelo razlikujejo.
Vam lahko služi: mitotično vreteno-Podenota β ima nekoliko nižjo molekulsko maso (med 35 in 36 kDa).
-Po drugi strani je γ podenota veliko manjša in ima približno molekulsko maso 8 kDa.
Vsi heterotrofimerni g. Ti dve domeni sta tako močno povezani, da se predvidevata kot enotna funkcionalna enota.
Fantje
Kot smo že omenili, obstajata dve vrsti G beljakovin: majhne in heterotrimérice.
Majhni G proteini imajo funkcije pri rasti celic, izločanju beljakovin in interakciji medceličnih veziklov. Heterotrofimerni G proteini so povezani s transdukcijo signala iz površinskih receptorjev in delujejo tudi kot stikala, ki se izmenjujejo med dvema stanjem, odvisno od povezanega nukleotida.
Majhni G proteini
Te beljakovine imenujemo tudi majhne GTPASA, majhne GTP stične beljakovine ali superdružine RAS in tvorijo neodvisno superdružino znotraj velikega razreda hidrolaze GTP z regulativnimi funkcijami.
Ti beljakovini so zelo raznoliki in nadzorujejo več celičnih procesov. Za njih je značilna ohranjena domena vezave na GTP, domena "g". Zveza tega nukleotidnega fosfata povzroči majhne konformacijske spremembe v svoji katalitični domeni v GD proteinih.
Njegova dejavnost je tesno povezana z beljakovinami, ki aktivirajo GTPASAS (GAP, angleščina "Protein, ki aktivira GTPase ") in z dejavniki izmenjave nukleotidov v Guanini (GEF, angleščina "Faktor izmenjave nukleotidnih nukleotidov ").
V evkariotih je bilo opisanih pet razredov ali družin majhnih G beljakovin:
-Ras
-Rho
-Rabi
-SAR1/ARF
-Tekel
RAS in Rho proteini nadzorujejo genetsko ekspresijo in Rho proteini tudi modulirajo reorganizacijo citoskeleta. Proteini skupine Rab in SAR1/ARF vplivajo na vezikularni transport, beljakovine pa uravnavajo jedrski transport in celični cikel.
Heterotrofimerni G proteini
Ta vrsta beljakovin si zasluži tudi povezavo z dvema drugim faktorjem beljakovin, tako da je signalna pot od zunaj do celične notranjosti sestavljena iz treh elementov v naslednjem vrstnem redu:
- The Povezani receptorji Beljakovine g
- The G
- The Beljakovine ali Učinkoviti kanali
Obstaja velika raznolikost heterotrofimernega G proteina in to je povezano z veliko raznolikostjo α podenot, ki obstajajo v naravi, v katerih se ohrani le 20% zaporedja aminokislin.
Običajno heterotrofimerni G proteini G se identificirajo zaradi raznolikosti α podenote, ki temelji predvsem na njihovi funkcionalni in zaporedni podobnosti.
Podenote α sestavljajo štiri družine (družina GS, družina GI/O, družina GQ in družina G12). Vsaka družina je sestavljena iz drugačnega "izotipa", ki skupaj doda več kot 15 različnih načinov α podenot.
Družina gs
Ta družina vsebuje predstavnike, ki sodelujejo tudi pri pozitivni regulaciji ciklaznih adenilatskih beljakovin in se izražajo v večini vrst celic. Sestavljen je iz dveh članov: GS in golf.
Naročnina "S " Nanaša se na stimulacijo (iz angleščine "Spodbuda ") in naročnina "Olf " Nanaša se na "vonj" (iz angleščine "Olfaction"). Golf beljakovine so izražene zlasti v senzoričnih nevronih, ki so odgovorni za vonj.
Družina gi/o
To je največja in raznolika družina. Izraženi so v številnih vrstah celic in posredujejo inhibicijo več vrst adenilciklase (podpisovanje "Yo " se nanaša na inhibicijo).
Beljakovine s skupinsko α podenotami se izražajo zlasti v celicah centralnega živčnega sistema in imajo dve različici: A in B.
Družina gq
Beljakovine s to družino α podenote so odgovorne za regulacijo fosfolipaze C. To družino sestavljajo štirje člani, katerih α podenote se izražajo z različnimi geni. V jetrnih celicah, v ledvičnih celicah in pljučih jih je obilo.
Družina g12
Ta družina je v organizmih vseprisotno izražena in z gotovostjo ni znano, kaj natančno so celični procesi, ki jih regulirajo z beljakovinami s temi podenotami.
Β in γ podenote
Čeprav je raznolikost alfa struktur odločilna za identifikacijo heterotimernih beljakovin, obstaja tudi veliko raznolikosti glede na druga dva podenota: beta in gama.
Funkcije
G proteini sodelujejo pri "kanalizaciji" signalov iz receptorjev v plazemski membrani do efektorskih kanalov ali encimov.
Najpogostejši primer funkcije te vrste beljakovin je pri uravnavanju encima adenilat ciklasa, encima, ki je odgovoren za sintezo adenozina 3 ', 5'-monofosfata ali preprosto cikličnega ojačevalnika, molekule, ki ima pomembne funkcije kot drugi glasnik kot drugi glasnik V mnogih znanih celičnih procesih:
Vam lahko služi: kaliciformne celice-Selektivna fosforilacija beljakovin s specifičnimi funkcijami
-Genetska transkripcija
-Reorganizacija citoskeleta
-Izločanje
-Membranska depolarizacija
Prav tako posredno sodelujejo pri regulaciji inozitolskega signalnega slapa (fosfatidilinitola in njihovih fosforiliranih derivatov), ki so odgovorni za nadzor procesov, odvisnih od kalcija, kot so kemiotaksija, in izločanje topnih faktorjev.
Številne ionske kanale in transportne beljakovine neposredno nadzirajo beljakovinski beljakovine g. Podobno so ti proteini med drugim vključeni v številne senzorične procese, kot so vid, vonj.
Kako delujejo?
Način interakcije G proteina z efektorskimi beljakovinami je značilen za vsak razred ali družino beljakovin.
Za G-grebranalni receptor G proteine (heterotrofimerne G proteine) združitev gvaninskega nukleotida, kot sta BDP ali Guanosín difosfat, do podenote α povzroči povezavo treh podenot, ki tvori kompleks, znan kot Gαβγ ali G-GDP, ki je povezana do membrane.
Če se molekula BDP pozneje izmenjuje z molekulo GTP, α podenota, pritrjena na GTP beljakovine.
Hidrolitna aktivnost te podenote vam omogoča, da končate aktivacijo, izmenjate GTP za nov BDP, premaknete se na neaktivno konformacijo.
Ker vzbujenih receptorjev, ki so povezani s postopkom izmenjave BDP BDP, je zelo počasen, kar pomeni, da heterotrimični GD proteini BDP izmenjujejo samo z GTP s fiziološko pomembno hitrostjo, ko se pridružijo svojim navdušenim receptorjem.
Reference
- Gilman, g. (1987). G proteini: pretvorniki signalov, ki jih vodi sprejemnik. Letna ocena biokemije, 56, 615-649.
- Milligan, g., & Kostenis in. (2006). Heterotrimični G-proteini: Kratka zgodovina. Britanski časopis za farmakologijo, 147, 546-555.
- Offermanns, s. (2003). G-proteini kot pretvorniki v transmembranski signalizaciji. Napredek v biofiziki in molekularni biologiji, 83, 101-130.
- Simon, m., Strathmann, m. Str., & Gautam, n. (1991). Raznolikost G proteinov pri transdukciji signala. Znanost, 252, 802-808.
- Syrovatka, v., Veselo, k. Tudi., Dey, r., & Huang, x. (2016). Regulacija, signalizacija in fiziološke funkcije G-proteinov. Časopis za molekularno biologijo, 428(19), 3850-3868.
- « Značilnosti Crotalus triseriatus, habitat, razmnoževanje
- Značilnosti groma, habitat, taksonomija, nepremičnine »