

Glikogenska struktura, sinteza, razgradnja, funkcije
- 2858
- 812
- Stuart Armstrong
On Glikogen To je ogljikovo hidrat večine sesalcev. Ogljikovi hidrati običajno imenujemo sladkor in jih razvrstimo glede na število odpadkov, ki jih povzročajo hidroliza (monosaharidi, disaharidi, oligosaharidi in polisaharidi)).
Monosaharidi so najpreprostejši ogljikovi hidrati, ki so razvrščeni glede na število ogljikov v njihovi strukturi. Takrat so triosi (3C), tetrosas (4C), pentosas (5C), heksoozni (6C), heptozaza (7C) in oktoze (8c).
Kemična struktura glikogena, ki prikazuje glikozidne vezi (vir: glikogen.SVG: Nevrotoger Derivatno delo: Marek M [javna domena] prek Wikimedia Commons) Glede na prisotnost aldehidske skupine ali Cetona skupine so ti monosaharidi razvrščeni tudi kot aldije oziroma ketosas.
Disaharidi povzročajo hidrolizo, dva preprosta monosaharida, medtem ko oligosaharidi proizvajajo 2 do 10 enot monosaharidov in polisaharidov, proizvajajo več kot 10 monosaharidov.
Glikogen je z biokemičnega vidika polisaharid, sestavljen iz razvejanih verig šestega ogljikovega aldoza, torej heksoza, znanega kot glukoza. Grafično ga je mogoče predstaviti za glikogen kot drevo glukoze. To se imenuje tudi živalski škrob.
Glukoza v rastlinah je shranjena kot škrob in pri živalih kot glikogen, ki je shranjen predvsem v jetrnem in mišičnem tkivu.
V jetrih lahko glikogen vzpostavi 10% svoje mase in 1% mišične mase. Kot pri 70kg moškemu jetra tehta približno 1800 g, mišice pa približno 35 kg, je skupna količina mišičnega glikogena veliko večja od jeter.
[TOC]
Struktura
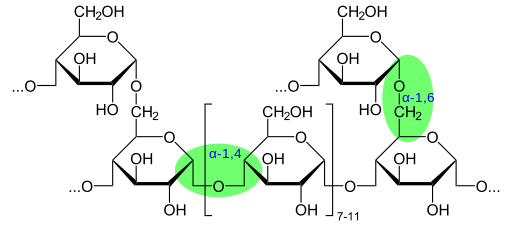
Molekulska masa glikogena lahko doseže 108 g/mol, kar ustreza molekuli glukoze 6 × 105. Glikogen je sestavljen iz več razvejanih α-D-glikoznih verig. Glukoza (C6H12O6) je aldoheksosa, ki jo je mogoče predstaviti na linearni ali ciklični način.
Glikogen ima zelo razvejano in kompaktno strukturo z verigami od 12 do 14 glukoznih odpadkov v obliki α-D-glukoze, ki so povezani z α- (1 → 4) glukozidnimi vezmi. Pojavite verige tvorijo α- (1 → 6) glukozidne povezave.
Glikogen, tako kot škrob, ki ga zaužijemo v prehrani, zagotavlja večino ogljikovih hidratov, ki jih telo potrebuje. V črevesju se te polisaharide razgradijo s hidrolizo in nato absorbirajo proti krožnemu hudourniku predvsem kot glukoza.
Trije encimi: ß-amilaza, α-amilaza in amilo-α- (1 → 6)-glukozidaza so odgovorni za črevesno razgradnjo tako glikogena kot škroba.
Α-amilaza naključno hidrolizira α-vezi (1 → 4) stranskih verig tako glikogena kot škroba, zato prejme ime endoglizidaze. Ss-Amila je eksoglikozidaza, ki sprošča ß-maltose dímeros, ki lomi α- (1 → 4) glikozidne povezave s koncev najbolj zunanjih verig, ne da bi dosegli posledice posledice.
Glede na dejstvo, da niti ß-amilaza niti α-amilaza razgradi veje, je končni produkt njegovega delovanja zelo razvejana struktura približno 35 do 40 ostankov glukoze, ki se imenujejo mej.
Mejna dekstrin se končno hidrolizira na točkah vej, ki imajo α- (1 → 6) vezi skozi Amyle-α- (1 → 6)-glukozidaza, znana tudi kot "obrekovalni" encim. Verige, ki jih sprošča ta defloat.
Ker zaužit glikogen vstopi kot glukoza, mora organizem sintetizirati v tkivih iz glukoze.
Vam lahko služi: purini: značilnosti, struktura, funkcijeSinteza
Sinteza glikogena se imenuje glikogeneza in poteka zlasti v mišicah in jetrih. Glukoza, ki v organizem vstopi s prehrano.
Glukokinaza fosforilna do glukoze v ogljiku 6. ATP zagotavlja fosfor in energijo za to reakcijo. Kot rezultat tega se tvori glukozni 6-fosfat in sprosti ADP. Nato 6-fosfatna glukoza postane glukoza 1-fosfat z delovanjem fosfoglukomutaze, ki blata fosfor iz položaja 6 v položaj 1.
1-fosfatna glukoza se aktivira za sintezo glikogena, kar pomeni udeležbo nabora treh drugih encimov: UDP-glikozno pirofosforilazo, sintetični glikogen in amilo- (1,4 → 1.6) -glikosiltransferaza.
Glukoza-1-fosfat, skupaj z trifosfatnim uridinom (UTP, nukleozidom uridin trifosfata) in z delovanjem UDP-glikoze-pirofosforilaze tvorijo dihosfat-glukozni uridinski kompleks (UDP GLC) (UDP GLC). V procesu je pirofosfatni ion hidroliziran.
Nato sintetiran encim glikogena tvori glukozidno vez med C1 UDP GLC kompleksa in C4 glikogenskega terminala glukoze, UDP pa se sprosti iz UDP aktiviranega glukoznega kompleksa. Da bi prišlo do te reakcije, mora obstajati že obstoječa molekula glikogena, imenovana "primarni glikogen".
Primordialni glikogen je sintetiziran na beljakovini, glikogenin, ki ima 37 kDa in glizila v ostanku tirozina z uporabo UDP GLC kompleksa. Od tam so povezani α-D-glukozni odpadki z 1 → 4 povezave in nastane majhna veriga, na kateri deluje sintetični glikogen.
Ko začetna veriga poveže vsaj 11 ostankov glukoze, encim razvejane ali amile- (1,4 → 1,6) -glikoziltransferaza prenaša verižni kos 6 ali 7 glukoze v sosednjo verigo v položaju 1 → 6, ki vzpostavi vejo točka. Tako zgrajena molekula glikogena raste z dodatki glukoznih enot z glikozidnimi povezavami 1 → 4 in več posledic.
Degradacija
Razgradnja glikogena se imenuje glukogenoliza in ni enakovredna povratni poti njegove sinteze. Hitrost te poti je omejena s hitrostjo reakcije, ki jo katalizira glikogen fosforilaze.
Glikogen fosforyllaze je odgovoren za cepitev (fosforoliza) povezav 1 → 4 iz glikogenskih verig, sproščanje glukoze 1-fosfat. Encimsko delovanje se začne na koncih najbolj oddaljenih verig in se odstrani zaporedno, dokler 4 ostanki glukoze ostanejo na vsaki strani posledic.
Nato še en encim, α- (1 → 4) → α- (1 → 4) Glucano transferas pusti vejsko točko, ki je izpostavljena s prenosom trisaharidne enote iz ene veje v drugo. To omogoča hidrolije amilo- (1 → 6) -glukozidaza (nerazrešeni encim). Kombinirano delovanje teh encimov se konča v celoti cepi na glikogen.
Ker je začetna reakcija fosfomutaze reverzibilna, lahko 6-fosfatna glukoza tvorimo iz ostankov glukoze 1-fosfat, razdeljen iz glikogena. V jetrih in ledvicah, vendar ne v mišici, je encim, glukoza-6-fosfataza, ki se lahko zbere na 6-fosfatno glukozo in jo spremeni v prosto glukozo.
Vam lahko služi: fotolizaDefinirana glukoza se lahko razširi na kri in tako se jet jetrna glikogenoliza odraža pri povečanju vrednosti glukoze v krvi (glikemija).
Regulacija sinteze in razgradnje
Sinteze
Ta postopek se izvaja na dveh temeljnih encimih: glikogenu s sintezaznim glikogenom in fosforilazo, tako da je, ko je eden od njih aktiviran, drugi v svojem neaktivnem stanju. Ta regulacija preprečuje, da bi se hkrati pojavile nasprotne reakcije sinteze in razgradnje.
Aktivna oblika in neaktivna oblika obeh encimov je zelo različna, medsebojna pretvorba aktivnih in neaktivnih oblik fosforilaze in sintetičnega glikogena.
Adrenalin je hormon, ki se sprošča iz nadledvičnega mozga, Glucagon. Endokrina trebušna slinavka proizvaja inzulin in glukagon. Langerhansovi otočki α so tisti, ki sintetizirajo glukagon.
Adrenalin in glukagon sta dva hormona, ki se sprostita, ko je potrebna energija kot odgovor na znižanje ravni glukoze v krvi. Ti hormoni stimulirajo aktivacijo fosforilaze glikogena in zavirajo sintetični glikogen, s čimer stimulirajo glikogenolizo in zavirajo glikogenezo.
Medtem ko adrenalin izvaja svoje delovanje na mišicah in jetrih, glukagon deluje samo na jetra. Ti hormoni so združeni s specifičnimi membranalnimi receptorji v beli celici, ki aktivira ciklasa adenilat.
Aktivacija ciklaze adenilata začne encimski slap, ki na eni strani aktivira AMPC -odvisno proteinkinazo, ki neaktivno za sintetični glikogen, in aktivira glikogeno fosforilazo s fosforilacijo (neposredno in indirktivno).
Skeletna mišica ima še en mehanizem za aktiviranje fosforilaze glikogena skozi kalcij, ki se sprošča kot posledica depolarizacije mišične membrane na začetku krčenja.
Degradacije
Zgoraj opisani encimski slapi na koncu povečajo raven glukoze in ko dosežejo določeno raven, se aktivira glikogeneza in zavira glukogenoliza, kar zavira tudi nadaljnje sproščanje adrenalina in glukagona.
Glikogenezo se aktivira z aktiviranjem fosfatazne fosforilaze, encima, ki uravnava sintezo glikogena z več mehanizmi, kar pomeni inaktivacijo kinaze fosforilaze in fosforilaze α, kar je sintetična zaviralec glikogena glikogena.
Inzulin spodbuja vnos glukoze v mišične celice in poveča 6-fosfatno raven glukoze, kar spodbuja defiliranje in aktiviranje sintezaznega glikogena. Tako se začne sinteza in razgradnja glikogena zavira.
Funkcije
Mišični glikogen predstavlja rezervat energije za mišico, ki tako kot rezervne maščobe omogoča mišici, da izpolni svoje funkcije. Ker je vir glukoze, se med vadbo uporablja mišični glikogen. Te rezervacije se s fizičnim treningom povečujejo.
V jetrih glikogen predstavlja tudi pomemben rezervni vir tako za funkcije organa kot za prispevek glukoze k preostalem telesu.
Ta funkcija jetrnega glikogena je posledica dejstva, da jetra vsebujejo 6-fosfatazno glukozo, encim, ki lahko izloči 6-fosfatno glukozno fosfatno skupino in jo spremeni v prosto glukozo. Prosta glukoza, za razliko od fosforilirane glukoze, se lahko širi po membrani hepatocitov (jetrne celice).
Lahko vam služi: sporulacija: v rastlinah, v glivah in v bakterijahTako lahko jetra zagotavljajo glukozo za obtok in vzdržujejo stabilno raven glukoze, tudi v dolgotrajnih pogojih na tešče.
Ta funkcija je zelo pomembna, saj se možgani negujejo skoraj izključno iz glukoze v krvi, zato lahko huda hipoglikemija (zelo nizke koncentracije glukoze v krvi) povzroči izgubo znanja.
Sorodne bolezni
Bolezni, povezane z glikogenom, prejmejo generično ime "bolezni shranjevanja glikogena".
Te bolezni predstavljajo skupino dednih patologij, za katero je značilno nahajališče v tkivih nenormalnih količin ali vrst glikogena.
Večino bolezni za shranjevanje glikogena povzročajo primanjkljaj genetske narave katerega koli od encimov, ki sodelujejo pri presnovi glikogena.
Uvrščeni so v osem vrst, večina pa ima svoja imena in vsako od njih proizvaja drugačen encimski primanjkljaj. Nekateri so smrtni v zelo zgodnjih življenjskih fazah, druge pa med vadbo spremljata mišična šibkost in primanjkljaj.
Izjemni primeri
Nekatere najpomembnejše bolezni, povezane z glikogenom, so naslednje:
- Bolezen Von Gierke ali bolezen shranjevanja glikogena tipa I, proizvaja 6-fosfatazni primanjkljaj glukoze v jetrih in ledvicah.
Zanj je značilna nenormalna rast jeter (hepatomegalija) zaradi pretiranega kopičenja glikogena in hipoglikemije, saj jetra ne morejo zagotoviti glukoze za obtok. Bolniki s tem stanjem imajo spremembe rasti.
- Bolezen Pompe ali tipa II je posledica α-primanjkljaja (1 → 4) -Glucano 6-glikoziltransferas v jetrih, srcu in skeletnih mišicah. Ta bolezen, kot sta Andersen ali Type IV, je smrtonosna pred dvema letoma življenja.
- Bolezen mcardle ali tipa V ima primanjkljaj mišične fosforilaze in ga spremljajo mišična šibkost, zmanjšana toleranca vadbe, nenormalno kopičenje mišičnega glikogena in odsotnost laktata med vadbo med vadbo.
Reference
- Bhattacharya, k. (2015). Preiskava in upravljanje bolezni shranjevanja glikogena. Translacijska pediatrija, 4(3), 240–248.
- Dagli, a., Sentner, c., & Weinstein, D. (2016). Glikogen shranjevanje bolezni tipa III. GENE MENCE, 1-16.
- Guyton, a., & Hall, J. (2006). Učbenik medicinske fiziologije (11. izd.). Elsevier Inc.
- Mathews, c., Van Holde, k., & Ahern, k. (2000). Biokemija (3. izd.). San Francisco, Kalifornija: Pearson.
- McKiernan, str. (2017). Patobiologija želje shranjevanja glikogena. Curr patobiol rep.
- Murray, r., Bender, d., Botham, k., Kennelly, str., Rodwell, v., & Weil, P. (2009). Harperjeva ilustrirana biokemija (28. izd.). McGraw-Hill Medical.
- Nelson, d. L., & Cox, m. M. (2009). Lehningerjeva načela biokemije. Omega izdaje (5. izd.).
- Rawn, j. D. (1998). Biokemija. Burlington, Massachusetts: Založniki Neil Patterson.
- Tarnopolsky, m. Do. (2018). Miopatije, povezane z motnjami metabolizma glikogena. Nevroterapevtika.
- « Argonska zgodovina, struktura, lastnosti, uporabe
- Bijektivna funkcija Kaj je, kako je, primeri, vaje »