

Oksidativne faze fosforilacije, funkcije in zaviralci
- 4587
- 97
- Raymond Moen
The Oksidativna fosforilacija Gre za postopek, pri katerem se molekule ATP sintetizirajo iz ADP in PYo (Anorganski fosfat). Ta mehanizem izvajajo bakterije in evkariontske celice. V evkariontskih celicah se fosforilacija izvaja v mitohondrijski matrici nefotosintetskih celic.
Proizvodnja ATP je usmerjena s prenosom elektronov iz koencimov NADH ali FADH2 do o2. Ta postopek predstavlja največjo proizvodnjo energije v celici in izhaja iz razgradnje ogljikovih hidratov in maščob.
Vir: robot8a [cc by-sa 4.0 (https: // creativeCommons.Org/licence/by-sa/4.0)] Energija, shranjena v gradientih obremenitve in pH, znana tudi kot protonska motorična sila, omogoča izvedbo tega postopka. Protonski gradient, ki nastane, povzroči, da ima zunanji del membrane pozitivno obremenitev zaradi koncentracije protonov (H+) in mitohondrijska matrika je negativna.
[TOC]
Kjer pride do oksidativne fosforilacije?
Procesi prenosa elektronov in oksidativne fosforilacije so povezani z membrano. V prokariotih se ti mehanizmi izvajajo skozi plazemsko membrano. V evkariontskih celicah so povezani z membrano mitohondrijev.
Število mitohondrijev, ki jih najdemo v celicah, se razlikuje glede na vrsto celice. Na primer, pri sesalcih eritrociti nimajo teh organelov, medtem ko imajo lahko druge vrste celic, kot so mišične celice.
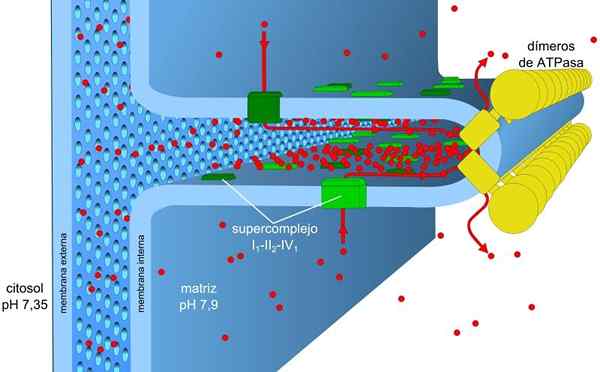
Mitohondrijska membrana je sestavljena iz preproste zunanje membrane, nekoliko bolj zapletene notranje membrane, sredi njih.
Zunanja membrana vsebuje beljakovine, imenovan porina, ki tvori kanale za preprosto difuzijo majhnih molekul. Ta membrana je odgovorna za vzdrževanje strukture in oblike mitohondrijev.
Notranja membrana ima večjo gostoto in je bogata z beljakovinami. Tudi za molekule in ione je vodoodporno, da jo prečkajo, potrebujejo intermembranalne beljakovine, ki jih prevažajo.
Znotraj matrice se raztezajo pregibi notranje membrane, ki tvorijo grebene, ki omogočajo, da ima veliko območje v majhni prostornini.
Celična energija centralna
Mitohondrije veljajo za celično energijsko središče. V njem so encimi, ki sodelujejo v procesih cikla citrične kisline, oksidacija maščobnih kislin in redoks encimov ter beljakovine transporta elektronov in doktorskega fosforilacije.
Koncentracijski gradient protonov (gradient pH) in gradient obremenitev ali električnega potenciala v notranji membrani mitohondrijev sta vzrok za prottonično motorično silo. Majhna prepustnost notranje membrane za ione (razen H+) omogoča, da imajo mitohondriji stabilen gradient napetosti.
Elektronski transport, črpanje protonov in ATP hkrati v mitohondrijih, zahvaljujoč protonski motorični sili. Gradient pH vzdržuje kisle razmere v intermembranski in mitohondrijski matrici z alkalnimi pogoji.
Za vsaka dva elektrona, prenesena v O2 Približno 10 protonov črpa membrana, kar ustvarja elektrokemični gradient. Energija, ki je v tem postopku, se postopoma proizvaja s prehodom elektronov skozi transportno verigo.
Obdobja
Energija, ki se sprošča med reakcijami oksida redukcije NADH in FADH2 Je znatno visok (približno 53 kcal/mol za vsak par elektronov), zato ga je treba uporabiti pri proizvodnji molekul ATP, postopoma ga je treba s prehodom elektronov skozi transporterje.
Ti so organizirani v štirih kompleksih, ki se nahajajo v notranji mitohondrijski membrani. Povezava teh reakcij na sintezo ATP se izvede v petem kompleksu.
Vam lahko služi: B limfociti: značilnosti, struktura, funkcije, vrsteTransportna veriga elektronov
NADH prenese nekaj elektronov, ki vstopijo v kompleks I transportne verige elektronov. Elektroni se prenesejo v mononukleotid Flavina in nato v ubikinon (koencim q) skozi železov transporter. Ta postopek sprosti veliko energije (16,6 kcal/mol).
Ubiquinona prevaža elektrone skozi membrano do kompleksa III. V tem kompleksu elektroni gredo skozi citokrome B in C1 Hvala transporterju železov.
Iz kompleksa III elektroni prehajajo v kompleks IV (citokrom c oksidaza), ki so ga prenesli s citokromom C (periferna membranska protein). V kompleksu IV elektroni prehajajo skozi par bakrenih ionov (CUdo2+), nato na citokrom cdo, Nato na drug par bakrenih ionov (Cub2+) in od tega do citokroma do3.
Končno se elektroni prenesejo v O2 ki je zadnji sprejemnik in tvori molekulo vode (h2O) Za vsak par prejetih elektronov. Prehod elektronov iz kompleksa IV v O2 Prav tako ustvarja veliko proste energije (25,8 kcal/mol).
Succinato Coq reduktaza
II kompleks (succinat coq reduktase) prejme par elektronov iz cikla citronske kisline zaradi oksidacije sukcinatne molekule do fumarata. Ti elektroni se prenesejo v FAD in nato skozi železno-azufre skupine, v Ubiquinona. Iz tega koencima gredo v kompleks III in sledijo prej opisani poti.
Energija, sproščena v reakciji prenosa elektronov na FAD, ni dovolj za povečanje protonov skozi membrano, tako da v tem koraku verige ni prottonične motorične sile, zato FADH daje manj H+ kot Nadh.
Energetska sklopka ali transdukcija
Energija, pridobljena v predhodno opisanem procesu prevoza elektronov, je treba uporabiti za proizvodnjo ATP, reakcijo, ki jo katalizira sintaza ATP ali kompleksni encim V. Ohranjanje takšne energije je znano kot priklop energije, mehanizem pa je bilo težko opisati.
Za opis te transdukcije energije je bilo opisanih več hipotez. Najbolje sprejeta je hipoteza kemosmotske spajanja, opisana spodaj.
Kemosmotska sklopka
Ta mehanizem predlaga, da energija, ki se uporablja za sintezo ATP, izvira iz protonskega gradienta v celičnih membranah. Ta postopek posega v mitohondrije, kloroplaste in bakterije in je povezan s prenosom elektronov.
I in IV kompleksi elektronskega transporta delujejo kot protonske črpalke. Te trpijo zaradi konformacijskih sprememb, ki jim omogočajo črpanje protonov v intermembranalni prostor. V kompleksu IV za vsak par elektronov se dva protona črpata zunaj membrane in še dva ostaneta v matrici, ki tvori H2Tudi.
Ubiquinona v kompleksu III sprejema protone kompleksov I in II in jih osvobodi zunaj membrane. I in III kompleksi omogočajo prehod štirih protonov za vsak par prevoženih elektronov.
Mitohondrijska matrika ima nizko koncentracijo protonov in negativnega električnega potenciala, medtem ko intermembranalni prostor predstavlja inverzne pogoje. Pretok protonov skozi to membrano pomeni elektrokemični gradient, ki shrani potrebno energijo (± 5 kcal/mol s protonom) za sintezo ATP.
Sinteza ATP
Encim ATP sintetaze je peti kompleks, ki sodeluje pri oksidativni fosforilaciji. Je odgovoren za izkoriščanje energije elektrokemičnega gradienta, da oblikuje ATP.
Ta transmembraal protein je sestavljen iz dveh komponent: F0 in f1. Komponenta f0 Omogoča vrnitev protonov v mitohondrijsko matrico, ki deluje kot kanal in F1 katalizira sintezo ATP prek ADP in PYo, z uporabo energije omenjenega donosa.
Vam lahko služi: membranski transportni proteini: funkcije in vrsteProces sinteze ATP zahteva strukturno spremembo F1 in sklop komponent F0 in f1. Translokacija protonov skozi F0 Povzroči konformacijske spremembe v treh podenomah F1, omogoča, da to deluje kot rotacijski motor, usmerjanje tvorbe ATP.
Podenota, ki je odgovorna za zvezo ADP s PYo prehaja iz šibkega stanja (l) v aktivno (t). Ko se oblikuje ATP, druga podenota preide v odprto stanje (o), ki omogoča sproščanje te molekule. Po izdaji ATP ta podenota prehaja iz odprtega stanja v neaktivno stanje (L).
Molekule ADP in PYo Pridružijo se podenoti, ki je prešla iz države ali države L.
Izdelki
Elektronska transportna veriga in fosforilacija proizvajata molekule ATP. Oksidacija NADH proizvede približno 52,12 kcal/mol (218 kJ/mol) proste energije.
Globalna reakcija za oksidacijo NADH je:
NADH+1⁄2 oz2 +H+ ↔ h2O+NAD+
Prenos elektronov iz NADH in FADH2 Pojavi se skozi več kompleksov, kar omogoča, da se sprememba proste energije ΔG ° razdeli na manjše "pakete", ki so povezani s sintezo ATP.
Oksidacija molekule NADH ustvari sintezo treh molekul ATP. Medtem ko oksidacija molekule FADH2 Je pritrjen na sintezo dveh ATP.
Ti koencimi izvirajo iz procesov glikolize in cikla citronske kisline. Za vsako razgrajeno molekulo glukoze se konča 36 ali 38 molekul ATP, odvisno od lokacije celic. V možganih in skeletnih mišicah se proizvaja 36 ATP, medtem ko se v mišičnem tkivu proizvaja 38 ATP.
Funkcije
Vsi organizmi, enocelični in večcelični, potrebujejo minimalno energijo v svojih celicah, da izvedejo procese znotraj njih, in posledično ohranjajo vitalne funkcije v celotnem telesu.
Metabolični procesi zahtevajo izvedbo energije. Večino uporabne energije dobimo z razgradnjo ogljikovih hidratov in maščob. Ta energija izhaja iz procesa oksidativne fosforilacije.
Nadzor oksidativne fosforilacije
Hitrost uporabe ATP v celicah nadzoruje sintezo istega, posledično pa zaradi spajanja oksidativne fosforilacije z verigo elektronov na splošno uravnava tudi elektronsko transportno hitrost.
Oksidativna fosforilacija ima strog nadzor, ki zagotavlja, da se ATP ne ustvarja hitreje, kot je porabljena. Obstajajo določeni koraki v procesu prenosa elektronov in povezane fosforilacije, ki uravnavajo stopnjo proizvodnje energije.
Koordinirani nadzor proizvodnje ATP
Glavne poti proizvodnje energije (celični ATP) so glikoliza, cikel citronske kisline in oksidativna fosforilacija. Koordinirani nadzor teh treh procesov uravnava sintezo ATP.
Nadzor fosforilacije z masnim delovanjem ATP je odvisen od natančnega prispevka elektronov v verigi transporte. To je odvisno od odnosa [NADH]/[NAD+] ki se ohrani povišano z delovanjem glikolize in cikla citronske kisline.
Ta usklajena kontrola se izvaja z uravnavanjem kontrolnih točk glikolize (citrat inhibira PFK) in cikel citrične kisline (dehidrogenaza piruvat, rezizan citrat, itchitrat dehidrogenaza in α-zetoglutarata dehidrogenaze).
Nadzor nad sprejemnikom
IV kompleks (citokrom c oksidaza) je encim, ki ga regulira eden od njegovih substratov, to je, da njegovo aktivnost nadzira zmanjšan citokrom C (C2+), kar je v ravnovesju z odnosom koncentracij med [NADH]/[NAD+] in razmerje masnega delovanja [ATP]/[ADP] + [PYo].
Lahko vam služi: ton: značilnosti in funkcijeVišji je [NADH]/[NAD+] in znižajte [ATP]/[ADP]+[PYo], več koncentracije bo citokrom [c2+] in večja bo dejavnost kompleksnega IV. To se na primer razlaga, če organizme primerjamo z različnimi dejavnostmi počitka in visokimi aktivnostmi.
Pri posamezniku z visoko telesno aktivnostjo poraba ATP in zato njena hidroliza na ADP + PYo To bo zelo visoko, kar bo povzročilo razliko v razmerju med masnimi dejanji, ki povzroči povečanje [c2+in zato povečanje sinteze ATP. Pri posamezniku, ki počiva, se pojavi obratna situacija.
Na koncu se hitrost oksidativne fosforilacije poveča s koncentracijo ADP znotraj mitohondrijev. Ta koncentracija je odvisna od translakatorjev ADP-ATP, zadolženih za transport adeninskih nukleotidov in PYo Od citosola do mitohondrijske matrice.
Sredstva za dekakoplantate
Na oksidativno fosforilacijo vplivajo nekatera kemična sredstva, ki omogočajo, da se elektronski transport nadaljuje brez fosforilacije ADP, ki loči proizvodnjo in ohranjanje energije.
Ta sredstva stimulirajo hitrost porabe kisika mitohondrijev v odsotnosti ADP, kar povzroči povečanje hidrolize ATP. Delujejo z odpravo posrednika ali razbijanjem energije iz verige elektronskih transportnih verig.
2,4-dinitrofenol, šibka kislina, ki poteka skozi mitohondrijske membrane, je odgovorna za razpršitev gradienta protonov, ker se jim pridružijo na kisli strani in jih sprostijo na osnovni strani.
Ta spojina je bila uporabljena kot "shujšava tableta", ker je bilo ugotovljeno, da je povečala dihanje, torej povečanje hitrosti in s tem povezano izgubo teže. Vendar se je pokazalo, da lahko njegov negativni učinek celo povzroči smrt.
Razprševanje protonskega gradienta proizvaja toploto. Rjave maščobne tkivne celice uporabljajo ločevanje, hormonalno nadzorovano, za proizvodnjo toplote. Hibernacijski sesalci in novorojenčki, ki nimajo las, so sestavljeni iz tega tkiva, ki jim služi kot nekakšna toplotna odeja.
Zaviralci
Zaviralne spojine ali povzročitelji preprečujejo porabo ali2 (elektronski transport), kot je povezana oksidativna fosforilacija. Ta sredstva se izogibajo tvorbi ATP z uporabo energije, proizvedene v elektronskem transportu. Zato se transportna veriga ustavi, saj takšna poraba energije ni urejena.
Oligomicinski antibiotik deluje kot zaviralec fosforilacije pri mnogih bakterijah, kar preprečuje stimulacijo ADP pred sintezo ATP.
Obstajajo tudi ionoforozna sredstva, ki sestavljajo topne maščobe s kationi, kot je K+ in na+, in prehodite mitohondrijsko membrano s temi kationi. Mitohondriji nato porabijo energijo, proizvedeno v elektronskem transportu za črpanje kationov, namesto da bi sintetizirali ATP.
Reference
- Alberts, b., Bray, d., Hopkin, k., Johnson, a., Lewis, J., Raff, m., Roberts, k. & Walter, str. (2004). Bistvena celična biologija. New York: Garland Science.
- Cooper, g. M., Hausman, r. In. & Wright, n. (2010). Celica. (str. 397-402). Marbán.
- Devlin, t. M. (1992). Učbenik biokemije: s kliničnimi korelacijami. John Wiley & Sons, Inc.
- Garrett, r. H., & Grisham, c. M. (2008). Biokemija. Thomson Brooks/Cole.
- Lodish, h., Darnell, J. In., Berk, a., Kaiser, c. Do., Krieger, m., Scott, m. Str., & Matsudaira, str. (2008). Biologija mollekulskih celic. Macmillan.
- Nelson, d. L., & Cox, m. M. (2006). Lehningerjeva načela biokemije 4. izdaja. Ed Omega. Barcelona.
- Voet, d., & Voet, J. G. (2006). Biokemija. Ed. Pan -american Medical.
- « Značilnosti euglenophyta, razmnoževanje, prehrana, klasifikacija
- Trening, funkcije in vrste/skupine glukozidov »