

Struktura enolaze, mehanizem delovanja, funkcije
- 3359
- 574
- Ricky Dach
The Enolaza To je encim, zadolžen za izvajanje pretvorbe D-2-fosfoglicerata (2pGA) v fosfoenolpiruvatu (PEP) v glikolizi in reverzni reakciji v glukoneogenezi, dve presnovni poti, ki sta del presnove celične energije.
Odločitev za katalizacijo te reakcije v eni ali drugi smeri je odvisna od dostopa, ki ga ima celica do glukoze. To je potreba po prilagajanju metabolizma na razgradnjo ali sintezo, da bi pridobila energijo. Nepogrešljivo za uresničitev njegovih vitalnih procesov.
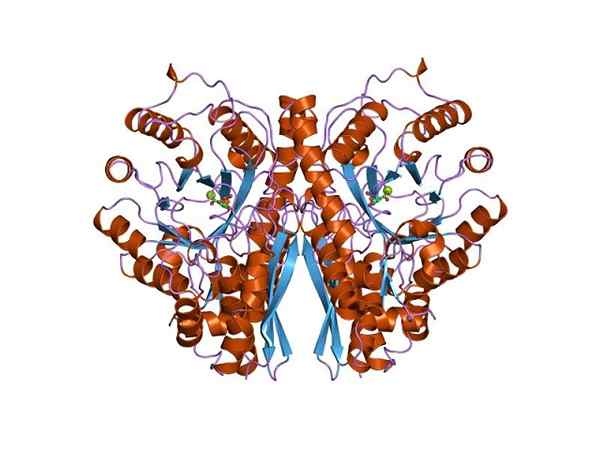
Tri -dimenzionalna struktura Enolasa. Avtor Jawahar Swaminathan in MSD osebje na Evropskem inštitutu za bioinformatiko [javna domena (https: // creativeCommons.Org/licence/by-sa/4.0)], iz Wikimedia Commons. Ker obe presnovni poti spadata v središče osrednjega metaboličnega drevesa živih bitij, ni čudno, da se aminokislinsko zaporedje tega proteina ohrani v Archeae, bakterijah in evkariotih. In zato predstavljajo podobne katalitične lastnosti.
Lokacija enolaze v celici je omejena na citosol, predal, v katerem potekata tako glikoliza (imenovana tudi glikoliza) in glukoneogeneza.
Vendar je bil odkrit tudi v drugih celičnih oddelkih, kot je plazemska membrana številnih rakavih celic in celic. Tam se zdi, da je vključen v olajšanje procesov razširjanja celic, kar je popolnoma drugačna funkcija od njegove klasične funkcije.
Encimi, ki so sposobni opravljati več funkcij, kot to počne Enolaza, so znani kot Encimi za lune.
[TOC]
Struktura
Kvarternarna struktura linusa ali ne v njegovih ligandih je bila določena v velikem številu prokariotskih in evkariotskih posameznikov.
Vsak monomer predstavlja dve domeni: majhno amino-terminalno domeno in večjo karboksilno-terminalno domeno. N-terminalna domena je sestavljena iz treh α in štirih β listov. Medtem ko je C-terminal sestavljen iz osmih β listov, ki se izmenjujejo med njimi, ki tvorijo β-sod, ki ga obdaja osem α propelerjev.
Poleg tega v vsakem monomeru najdemo dve sindikalni mesti za dvovalentne katione, ki se imenujeta "konformacijsko mesto" in "katalitično mesto". Prva ni zelo selektivna in se lahko pridruži najrazličnejšim dvovalentnim kationom, če ni substrata.
Vam lahko služi: sfingolipidi: kaj so, značilnosti, funkcije, sintezaMedtem ko se drugi veže na ione, potem ko se je substrat pridružil encimu. Zveza ionov na obeh mestih je ključnega pomena za nadaljevanje reakcije.
Končno je pomembno omeniti, da se v homodimerih monomerji pridružijo z ohranjanjem vzporedne orientacije. Zato je aktivno mesto omejeno na osrednjo regijo, ki jo oblikuje omenjena zveza.
Vendar v katalizi sodeluje le eden od obeh monomerov. To pojasnjuje, da sposobnost monomerov, da reakcijo izvedejo v eksperimentalnih pogojih.
Mehanizem delovanja
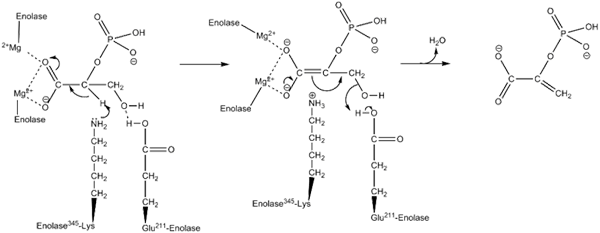 Mehanizem delovanja, ki ga uporablja encim inolaza. Avtor Kthompson08 v angleški Wikipediji [javna domena (https: // creativeCommons.Org/licence/by-sa/4.0)], iz Wikimedia Commons.
Mehanizem delovanja, ki ga uporablja encim inolaza. Avtor Kthompson08 v angleški Wikipediji [javna domena (https: // creativeCommons.Org/licence/by-sa/4.0)], iz Wikimedia Commons. Strukturne študije in tiste, ki so omogočile določitev kinetičnih in fizikalno -kemijskih značilnosti Enolasa, so nam omogočile razumevanje njihovega mehanizma delovanja.
Način, kako encim izvaja katalizo reakcije. Čeprav je vključen en substrat, je predlagan urejen zaporedni mehanizem.
To se začne z združenjem iona Mg2+ na konformacijsko mesto enega od monomerov. Neprekinjeno z zvezo substrata na aktivno mesto, ki ji sledi združitev drugega iona na katalitično mesto in zaključi s hitro sproščanjem izdelka, ko je reakcija izvedena. Na tej točki ostaja MG2+ povezan s konformacijskim mestom.
Po istih poteh daje prednost reakciji povprečnega encima na prvi stopnji nastajanja karbaniona posrednika, ki izloča proton ogljika 2 2PGA. To je zahvaljujoč delovanju osnovnega ostanka aminokisline.
Odstranjevanje hidroksila v ogljiku 3 poteka z delovanjem kislega ostanka iz encima. Na tej točki se zveza obeh ogljikov izvaja z dvojno vezjo, ki tvori PEP. Na ta način je reakcija vrhunec.
Lahko vam služi: Monera Kingdom: značilnosti, razvrstitev in primeriFunkcije
Številni doslej preučeni encimi so sposobni opravljati najrazličnejše funkcije, ki niso povezane z njihovo "klasično funkcijo" v različnih celičnih oddelkih. Ti encimi so bili imenovani "mesečni" encimi.
V tem smislu lahko Enolazo štejemo za encimsko luč, saj so bile številne funkcije, ki nasprotujejo njihovi klasični funkciji.
Nekatere od teh funkcij so naslednje:
- Sodelujte pri vzdrževanju celične oblike in vezikularnega prometa pri interakciji s citoskeletnimi beljakovinami.
- V jedru sesalskih celic deluje kot transkripcijski faktor, ki uravnava izražanje genov, povezanih s celično proliferacijo. Sodeluje pri vzdrževanju stabilnosti RNM v razgradbi v bakterijah.
- V patogenih, kot so Streptococcus pneumoniae in Trypanosoma cruzi, Zdi se, da deluje kot pomemben dejavnik virulence.
- Ugotovljeno je bilo tudi, da v Streptococcus pyogenes, Enolaza se izloči na zunajcelični medij, ki olajša razgradnjo tkiva in utaje gostiteljskega sistema.
- Na površini tumorskih celic se izraža z izboljšanjem metastaz.
Enolaza in njen odnos z mehanizmi razširjanja celic
Številni patogeni, pa tudi tumorske celice, izražajo v svoji membrani ali izločajo zunajcelične medijske proteaze, ki lahko razgradijo proteinske beljakovine.
Ta zmogljivost omogoča, da se te celice prebijejo skozi tkiva in se hitro širijo po celotnem gostiteljskem organizmu. Tako daje prednost utaji imunskega sistema in zato vzpostavitev okužbe.
Tudi če Enolasi nima proteaze, sodeluje v procesu razširjanja številnih patogenov v svojem gostitelju in tumorskih celicah med golom.
To doseže zaradi dejstva, da je na površini teh celic izražen kot plazminogenski receptor. Slednji je zimogen serin-protetaze, znane kot plazminat, ki je del fibrinolitičnega sistema, in deluje razgradnja zunajceličnih matričnih beljakovin.
Zato je enolaza, izražena na površini, strategija, ki so jo te celice pridobile za vzpostavitev okužbe in uspešno širjenje.
Lahko vam služi: simpatična specifikacija: koncept, značilnosti in primeriTa strategija je sestavljena iz dveh procesov:
- Utaje imunskega sistema gostitelja. Ker jih te celice iz gostiteljskega proteina prevlečejo, jih ne upoštevajo celice imunskega sistema, ki prepoznajo patogene, povezane s patogeni.
- Post-action razširjanje plazminogena v plazminiranju. Katerih sodelovanje pri razgradnji zunajceličnih matričnih beljakovin, nato olajša hitro in učinkovito razširjanje.
Reference
- AVILAN L, GUALDRON-LOPEZ M, QUIñones W, González-González L, Hannaert V, Michels Paa, Concepción Jl. Enolase: Ključni igralec v presnovi in verjeten faktor virulence tripanosomatidnih parazitov-perspektiv za njegovo uporabo kot terapevtski cilj. Encimske raziskave. 2011 Vol. Članek ID932549, 14 strani.
- BhowMick I, Kumar N, Sharma S, Coppens I, Jarori GK, Plasmodium falciparum Enosoase: stopnjo specifične ekspresije in podcelična lokalizacija. Malaria Journal. 2009; 8 (1). Člen 179.
- Dan I, Peshavaria M, Quinn GB, diferencialna molekularna ura v evoluciji Enoase izoproteina. Časopis za molekularno evolucijo. 1993; 36 (6): 599-601.
- od Torre-Scoder E, Manzano-Román R, Pérez-Sánchez R, Siles-Lucas M, Oleaga A. Kloniranje in karakterizacija površinske vezave plazminogena, ki je povezana Schistosoma Bovis. Veterinarska parazitologija. 2010; 173: 73–84.
- Dinovo EC, Boyer PD. Izotopske sonde reakcijskega mehanizma enolaze. Začetni in equiquibium izotopski tečaji: primarni in sekadarski učinki izotopov. J Biol Chem. 1971; 246 (14): 4586-4593.
- Kaberdin VR, Lin-Chao S, razkrivanje novih vlog za manjše komponente E. Coli RNA razgradnjo. RNA biologija. 2009; 6 (4): 402-405.
- Keller A, Peltzer J, Carpentier G. Interakcije Enolas. Biochimica et Biophysica Acta. 2007; 1770 (6): 919–926.
- Lung J, Liu KJ, Chang JY, Leu SJ, Shih NY. MBP-1 je učinkovito kodiran z alternativnim prepisom gena ENRI1, ki je reguliran s proteinskim premikom proteina. FEBS Journal. 2010; 277 (20): 4308-4321.
- Pancholi v. Večfunkcionalna α-enolaza: njegova vloga pri boleznih. Celične in molekularne življenjske vede. 2001; 58 (7): 902-920.
- Poyner RR, Cleland WW, Reed GH. Vloga kovinskih INS v katalizi z enolazo. Urejen kinetični mehanizem za en encim substrata. Biokemija. 2001; 40: 9008-8017.
- Segovia-Gamboa NC, Chávez-Munguía B, Medina-Flores a, Entamoeba Invadens, Proces encystation in enolaza. Eksperimentalna parazitologija. 2010; 125 (2): 63–69.
- Tanaka M, Sugisaki K, Nakashima K, preklapljanje ravni prevajalnih mRNA za enoase izocime med razvojem skeletnih mišic Chickn. Biokemična in biofizična raziskovalna komunikacija. 1985; 133 (3): 868-872.