

Citozinska struktura, funkcije, lastnosti, sinteza
- 2165
- 273
- Adrian Legros
The Citozin Gre za pirimidinsko dušikovo bazo, ki služi za biosintezo citidina-5'-monofosfata in deksicitidina-5'-monofosfata. Te spojine služijo za biosintezo iz deoksiribonukleinske kisline (DNK) in ribonukleinske kisline (RNA). DNK shrani genetske informacije in RNA ima različne funkcije.
Pri živih bitjih citozina ne najdemo prosto, ampak običajno ribonukleotidi ali deoksiribonukleotidi. Obe vrsti spojine imata fosfatno skupino, ribozo in bazo dušika.
Vir: Vesprcom [javna domena] Ogljik 2 ribose ima v ribonukleotidih skupino oksidrillo (-OH) in atom vodika (-H) v deoksiribonukleotidih. Glede na število prisotnih fosfatnih skupin obstajajo Citidín-5'-Monophosfat (CMP), Citidín-5'-difosfat (CDP) in Citidín-5'-Triffosfat (CTP) (CTP) (CTP).
Desoksigenirani ekvivalenti se imenujejo desoxicitidin-5'-monofosfat (DCMP), desoxicitidin-5'-difosfat (DCDP) in desoxicitidin-5'-Triffosfat (DCTP).
Citozin v različnih oblikah sodeluje v različnih funkcijah, kot so biosinteza DNK in RNA, biosinteza glikoproteinov in regulacija izražanja gena.
[TOC]
Struktura in lastnosti
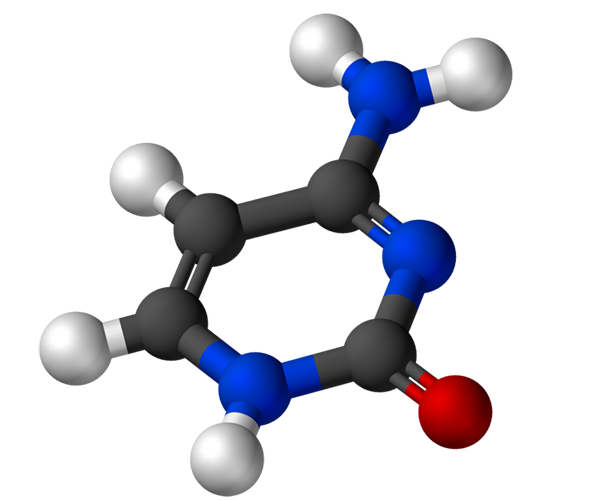
Citozin, 4-amino-2-hidroksipirimidin, ima empirično formulo C4H5N3Ali, katerih molekulska teža je 111,10 g/mol in je očiščena kot beli prah.
Struktura citozina je heterocikličen, aromatičen in ravninski obroč. Največja valovna dolžina absorbance (ʎMax) Je 260 nm. Fuzijska temperatura citozina presega 300 ° C.
Za tvorbo nukleotida se citozin kovalentno veže, prek dušika 1. Carbon 5 'je esterificiran s fosfatno skupino.
Biosinteza
Birimidinska nukleotidna biosinteza ima skupno pot, ki jo sestavljajo šesti koraki, ki jih katalizirajo encimi. Cesta se začne z biosintezo karbamoil-fosfata. V prokariotih obstaja samo en encim: karbamoil fosfat sintaza. To je odgovorno za sintezo pirimidinov in glutamina. V evkariotih obstajata karbamoil fosfat sintaza I in II, ki sta odgovorna za biosintezo glutamina in pirimidina.
Drugi korak je sestavljen iz tvorbe N-carbamoilaspartato iz ogljikovega fosfata in aspartata, reakcije, ki jo katalizira transpartat transcabamoalasa (ATCASA).
Tretji korak je sinteza L-dihidrotoata, ki povzroči zapiranje pirimidinskega obroča. Ta korak katalizira dihidrootaza.
Četrti korak je tvorba orotata, ki je redoks reakcija, ki jo katalizira dihidroorotat dehidrogenaza.
Peti korak je sestavljen iz tvorbe zlatata (OMP) z uporabo fosforibozil pirofosfata (PRPP) kot substrata, in fosforibozil orotat transferaza kot katalizator kot katalizator.
Šesti korak je tvorba uridilaato (uridin-5'-monophosfat, UMP), reakcije, ki jo katalizira OMP-dekarboksilaza.
Lahko vam služi: flora in favna iz HidalgaNaslednji koraki so sestavljeni iz fosforilacije UMP, ki ga katalizirajo kinaze, da tvorijo UTP, in prenos amino skupine iz glutamina v UTP, da tvori CTP.
Uredba o biosintezi
Pri sesalcih regulacija poteka na karbamoil fosfatni sintazi.
Carbamoil Syntasa II regulira negativne povratne informacije. Njeni regulatorji, UTP in PRPP, sta zaviralec in aktivator tega encima.
V tkivih, ki niso jeter, je karbamoil fosfat sintaza II edini vir karbamoil fosfata. Medtem ko je v jetrih, pod pogoji presežka amoniaka, karbamoil fosfat sintaza I proizvaja v mitohondrijih, karbamoil fosfat, ki se prevaža v citosol, od koder vstopi na pot biosinteze pirimidinov.
Druga regulacijska točka je omp-discarboxilaza, ki jo regulira konkurenčna inhibicija. Produkt njegove reakcije, UMP, tekmuje z OMP za vezavno mesto v OMP-Descarboxilasa.
Pirimidini, tako kot citozin, se reciklirajo
Recikliranje pirimidinov ima kot funkcija ponovna uporaba pirimidinov brez potrebe po novo biosintezi in se izogne razgradni poti. Reakcija recikliranja katalizira fosforibosiltransferaza pirimimidin. Splošna reakcija je naslednja:
Pirimidin + prpp -> nukleozid pirimidin 5 '-monofosfat + ppi
V vretenčarjih je v erihrocitih pirimimidin fosforibosiltransferaze pirimimidin. Pirimidinski substrat tega encima so Uracil, Timina in Orotato. Citozin je posredno recikliran iz uridin-5'-monofosfata.
Delovanje v biosintezi DNK
Med podvajanjem DNK se informacije, ki jih vsebuje DNK, kopirajo v DNK prek DNK polimeraze.
Biosinteza RNA potrebuje deoksinukleotidi trifosfat (DNTP), in sicer: deksitimidni trifosfat (DTTP), deksicitizen trifosfat (DCTP), de -alksiksiadenin trifar. Reakcija je:
(DNK)n odpadki + DNTP -> (DNK)N+1 odpadki + ppi
Hidroliza anorganskega pirofosfata (PPI) zagotavlja energijo za biosintezo RNA.
Delovanje pri stabilizaciji strukture DNK
V dvojnem DNUS Helixu je purina verige povezana s pirimidinom, nasprotne verige, z vodikovimi vezmi. Tako je citozin vedno povezan z gvaninom s tremi vodikovimi vezmi: adenin je povezan s Timinom z dvema vodikovih vezima.
Lahko vam služi: pomen biologijeVodikove vezi se porušijo, ko prečiščena izvorna DNK raztopina pri pH 7 podvržemo temperaturam, večje od 80 ° C. To povzroči, da dvojni DNA propeler tvori dve ločeni verigi. Ta postopek je znan kot denaturacija.
Temperatura, pri kateri je 50% DNK denaturirana, je znana kot temperatura taljenja (TM). Molekule DNK, katerih gvanin in citozin.
Zgoraj je eksperimentalni test, da večje število vodikovih vezi bolje stabilizira domače molekule DNK.
Funkcija bogatih regij v citozinu v DNK
Nedavno je bilo ugotovljeno, da lahko DNK človeškega celičnega jedra sprejme interkalirane strukture motivov (IM). Te strukture se proizvajajo v citozinskih regijah.
Struktura IM je sestavljena iz štirih verig DNK, za razliko od klasične DNK dvojnega propelerja, ki ima dve verigi. Natančneje, dve vzporedni dvostranski verigi sta prepleteni v orientaciji antiparallela in jih vzdržujeta par hemiprotoniranih citozinov (c: c: c+).
V človeškem genomu se strukture IM najdejo v regijah, kot so promotorji in telomeri. Število struktur IM je večje med fazo G1/S celičnega cikla, pri kateri je transkripcija visoka. Te regije so mesta za prepoznavanje beljakovin, vključena v aktiviranje transkripcijskih strojev.
Po drugi strani pa DNK v regijah, ki so bogate z gvaninskimi bazami (c) zaporedni. Ta oblika je značilna za mešane pasove RNA in dvojne DNA-ARN med transkripcijo in podvajanjem, v določenih trenutkih, ko je DNK povezana z beljakovinami.
Pokazalo se je, da regije s zaporednimi citozinskimi bazami ustvarjajo elektropozitiven obliž v glavnem razcepju DNK. Zato se verjame, da se ta območja vežejo na beljakovine, kar nagne na nekatere genomske regije genetske krhkosti.
Delovanje v biosintezi RNA
Med prepisom se podatki, ki jih vsebuje DNK, kopirajo v RNA prek RNA Pon polimeraze. Biosinteza RNA potrebuje triposfatne nukleozide (NTP), in sicer: Tryphyse citidin (CTP), triposfat uridin (UTP), adenin triposfat (ATP) in trifosfat gvanin (GTP). Reakcija je:
Vam lahko služi: flora in favna iz Campecheja: reprezentativna vrsta(RNA)n odpadki + NTP -> (RNA)N+1 odpadki + ppi
Hidroliza anorganskega pirofosfata (PPI) zagotavlja energijo za biosintezo RNA.
Delovanje v biosintezi glikoproteinov
Zaporedni prenos heksoz, ki tvorijo oligosaharide, O-ligate na beljakovine, se zgodi iz nukleotidnih prekurzorjev.
V vretenčarjih je zadnji korak biosinteze oligosaharidov, ligiranih O. Ta reakcija nastane v trans vreči Golgi.
Zdravljenja citozina in kemoterapije proti raku
Tetrahidrofolatna kislina (FH4) je skupinski vir3, In je potrebno za biosintezo DTMP iz smetišča. Tudi FH2 je oblikovan. Zmanjšanje FH2 na FH4 zahteva folat in NADPH reduktazo. Pri zdravljenju raka se uporabljajo nekateri zaviralci folatne reduktaze, kot sta aminopterin in metretreksat.
Metotrekxan je konkurenčen zaviralec. Folat reduktaza se pridruži s 100 -krat večjo afiniteto do tega zaviralca kot na njegov substrat. Aminapterin deluje podobno.
Inhibicija folata reduktaze posredno ovira biosintezo DTMP in torej DCTP. Neposredna inhibicija se zgodi prek zaviralcev timidilatne sintetize, ki katalizira DTMP iz smetišča. Ti zaviralci so 5-fluorouracil in 5-fluoro-2-dioksiuridin.
Na primer, 5-fluoroacil ni zaviralec, ampak najprej postane pot recikliranja, v deoksiuridnem mofosfatu D (fdump), ki se veže na timidilatno sintezo in ga zavira.
Snovi, analogne glutaminu, azaserinu in acivicinu, zavirajo amidotransferazo glutamin. Azarin je bil ena prvih odkritih snovi, ki delujejo kot samomorilno inaktiviranje.
Reference
- ASSI, h.Do., Garavís, m., González, c., In Damha, m.J. 2018. I-motif DNK: strukturne značilnosti in pomen za celično biologijo. Nuclei Acid Research, 46: 8038-8056.
- Bohinski, r. 1991. Biokemija. Addison-Wesley Iberoamericana, Wilmington, Delaware.
- Devlin, t.M. 2000. Biokemija. Uredništvo Reverté, Barcelona.
- Lodish, h., Berk, a., Zipurski, s.L., Matsudaria, str., Baltimore, d., Darnell, J. 2003. Celična in molekularna biologija. Uredništvo Medica Panamericana, Buenos Aires, Bogota, Caracas, Madrid, Mehika, Sāo Paulo.
- Nelson, d. L., Cox, m. M. 2008. Lehning-principles biokemije. W.H. Freeman, New York.
- Voet, d. in voet, j. 2004. Biokemija. John Wiley in sinovi, ZDA.