

Kardiolipinska struktura, sinteza, funkcije
- 4218
- 29
- Barry Ernser
The Kardiolipin, Znan tudi kot defatidilglicerol, je lipid družine gliceofosfolipidov in skupine poliglifosfolipidov. Najdemo ga v mitohondrijski membrani evkariontskih organizmov, v plazemski membrani številnih bakterij in tudi v nekaterih lokih.
Pangborn ga je odkril leta 1942 iz analize membranskih lipidov srčnega tkiva goveda. Njegova struktura je bila predlagana leta 1956, kemična sinteza pa je potekala približno 10 let pozneje.
Struktura kardiolipina (vir: EDGAR181 [javna domena] prek Wikimedia Commons) Nekateri avtorji menijo, da je njihova prisotnost omejena na membrane, ki proizvajajo ATP, prav tako pa tudi mitohondrije v evkariotih, plazemske membrane v bakterijah in hidrogenosomih (tipa-mmitokondrije) v določenih prostistih).
Dejstvo, da je kardiolipin v mitohondrijih in v plazemski membrani bakterij, je bilo uporabljeno za okrepitev temeljev endosimbiotske teorije, ki ugotovi, da so mitohondrije nastale v potomskih celicah evkariote s fagocitozo bakterije, ki je potem postal odvisen celica in obratno.
Njegova biosintetska pot pri živalih je bila opisana med letoma 1970 in 1972, nato pa se je pokazalo, da gre za isto pot, ki se pojavlja v rastlinah, kvasovkah, glivah in nevretenčarjih. Ni zelo obilen lipid, vendar celice zahtevajo, da pravilno deluje.
Pomen tega fosfolipida za mitohondrije in torej za presnovo celic je razviden, ko okvarjeno delovanje presnovnih poti, povezane z njim.
[TOC]
Struktura
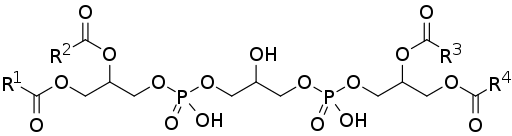
Kardiolipin ali deficientilglicerol je sestavljen iz dveh fosfatidnih kisline (lažje fosfolipide), povezanih med seboj prek molekule glicerola.
Fosfatidna kislina, eden od običajnih posrednikov na biosintetskih poteh drugih fosfolipidov, je sestavljena iz 3-fosfatne glicerolske molekule, ki sta na položajih karbonov 1 in 2, tako da sta dve verigi maščobnih kislin, zato je znan tudi kot 1,2 -Diacilglicerol 3-fosfat.
Vam lahko služi: sekundarna struktura beljakovin: značilnostiZato je kardiolipin sestavljen iz treh molekul glicerola: centralni glicerol, skupaj s fosfatno skupino v ogljiku 1, drugo fosfatno skupino v ogljiku 3 in hidroksilno skupino v ogljiku 2; in dva "stranska" glicerola.
Dve "stranski" glicerol molekuli se vežejo na osrednjo molekulo z "glicerol mostovi" skozi svoje ogljikove v položaju 3. V ogljikih položajev 1 in 2 sta esterificirali dve verigi maščobnih kislin in spremenljive nasičenosti.
Kardiolipin je lipid, ki lahko ali ne tvori bikapov, odvisno od prisotnosti ali odsotnosti dvovalentnih kationov. To je povezano z dejstvom, da gre za simetrično molekulo, zaradi česar je pomembna pri membranah, ki so odgovorni za procese transdukcije energije.
Tako kot drugi lipidi skupine poliglifosfolipidov ima tudi kardiolipin več hidroksilnih skupin, ki lahko služijo združitvi maščobnih kislin. Zato ima več pozicijskih stereoizomerov.
Njegove maščobne kisline
Različne študije so ugotovile, da so maščobne kisline, pritrjene na stranske molekule glicerola kardiolipina, običajno nenasičene, vendar stopnja nenasičenosti ni bila določena.
Nato imajo lahko takšne maščobne kisline med 14 in 22 ogljikovih dolžin in od 0 do 6 dvojnih vezi. To in dejstvo, da ima kardiolipin štiri vezane maščobne molekule, pomeni, da lahko obstaja več spremenljivk in kombinacije tega fosfolipida.
Sinteza
Biosinteza kardiolipina, kot je mogoče pričakovati, se začne s sintezo fosfatidne kisline ali 1,2-diacilglicerol 3-fosfata iz 3-fosfatnega glicerola in maščobnih kislin in maščobnih kislin. Ta postopek se pojavlja v mitohondrijih evkariotov in v plazemski membrani bakterij.
Vam lahko služi: ciklasa adenilate: značilnosti, vrste, funkcijeSinteza v evkariotih
Po tvorbi fosfatidna kislina reagira z visokoenergijsko molekulo, analogno ATP: CTP. Nato je posrednik, tudi z visoko energijo, znan kot fosfatidil-CMP. Aktivirana skupina fosfatidil se prenese v hidroksilno skupino v položaju C1 centralne molekule glicerola, ki služi kot okostje.
Ta proces rezultat. Ta prejme še eno fosfatidilno skupino, aktivirano iz druge fosfatidil-CMP molekule, reakcije, ki jo katalizira fosfatidiltransferaza, znana tudi kot kardiolipinska sintaza.
Encim s sintazo kardiolipin leži v notranji mitohondrijski membrani in zdi se, da tvori odličen kompleks, vsaj v kvasovkah. Njegov gen je izražen v velikih količinah v bogatih tkaninah v mitohondrijih, kot so srce, jetra in skeletna mišica vretenčarjev.
Ureditev njegove aktivnosti je v veliki meri odvisna od istih transkripcijskih faktorjev in endokrinih dejavnikov, ki modulirajo mitohondrijsko biogenezo.
Ko je sintetiziran v notranji mitohondrijski membrani, je treba kardiolipin premestiti v zunanjo mitohondrijsko membrano, tako da je podana vrsta topoloških procesov v membrani in drugi strukturni elementi iste.
Sinteza v prokariotih
Vsebnost kardiolipina v bakterijah je lahko zelo spremenljiva in je odvisna predvsem od fiziološkega stanja celic: v eksponentni fazi rasti je običajno manj obilna in bolj obilna, kadar se zmanjša (na primer v stacionarni fazi).
Njegovo biosintetsko pot lahko streljajo z različnimi stresnimi dražljaji, kot sta energetski primanjkljaj ali osmotski stres.
Dokler nastane fosfatidilglicerol, je postopek v evkariotih in prokariotih enakovreden, v prokariotih. To reakcijo katalizira encim fosfolipaza D, znano tudi kot kardiolipin sintaza.
Lahko vam služi: sglt (natrijeva glukoza transportna beljakovine)Ta reakcija je znana kot reakcija "transfatidila" (iz angleščine "Transposfatidilation "), Kjer eden od fosfatidilglicerolov deluje kot darovalec fosfatidilske skupine, drugi pa kot sprejemnik.
Funkcije
Fizične značilnosti kardiolipinskih molekul očitno omogočajo določene interakcije, ki opravljajo pomembne funkcije v strukturni organizaciji membran, kjer jih najdemo.
Med temi funkcijami so med drugim diskriminacija nekaterih membranskih domen, interakcije ali "navzkrižno povezanost" s transmarketnimi beljakovinami ali njihovimi poddomenami.
Zahvaljujoč svojim fizikalno -kemijskim značilnostim je kardiolipin prepoznan kot lipid, ki ne tvori Bápara, vendar lahko funkcija stabilizacija in "namestitev" transmembranskih beljakovin v lipidnih bicapah.
Zlasti njegove električne značilnosti dajejo funkcije v postopkih prenosa protona, ki potekajo v mitohondrijih.
Čeprav celice lahko preživijo brez tega fosfolipida, so nekatere študije ugotovile, da je potrebno za optimalno delovanje le -teh.
Reference
- Harayama, t., & Riezman, h. (2018). Razumevanje raznolikosti membranske lipidne sestave. Nature pregleduje molekularno celično biologijo, 19(5), 281–296.
- Luckey, m. (2008). Biološka strukturna membrana: z biokemičnimi in biofizičnimi temelji. Cambridge University Press.
- Murray, r., Bender, d., Botham, k., Kennelly, str., Rodwell, v., & Weil, P. (2009). Harperjeva ilustrirana biokemija (28. izd.). McGraw-Hill Medical.
- Van Meer, g., Voelker, d. R., & Feigenson, g. W. (2008). Membranski lipidi: kje so in kako se obnašajo. Ocene narave, 9, 112-124.
- Vance, j. In., & Vance, D. In. (2008). Biokemija lipidov, lipoproteinov in membran. V Novo razumevanje biokemije Vol. 36 (4. izd.). Elsevier.
- « Citozinska struktura, funkcije, lastnosti, sinteza
- Diacilglicerol struktura, biosinteza, funkcije »