

SGLT (natrijeva glukozna transportna beljakovine)
- 4644
- 624
- Dexter Koch
The Natrijeva-glukozna transportna beljakovine (SGLT) Odgovorni so za izvajanje aktivnega transporta glukoze v celicah sesalcev proti koncentracijskemu gradientu. Energija, potrebna za ta prevoz, se pridobiva od natrijevega kotransport v isti smeri (Simport).
Njegova lokacija je omejena na membrano celic, ki tvorijo epitelijska tkiva, ki so odgovorna za absorpcijo in reabsorpcijo hranil (tanko črevo in proksimalni konturirani tubul ledvice).
Do danes je bilo opisanih le šest izoform, ki pripadajo tej družini prevoznikov: SGLT-1, SGLT-2, SGLT-3, SGLT-4, SGLT-5 in SGLT-6. V vseh njih elektrokemični tok, ki ga ustvarja transport natrijevega iona, zagotavlja energijo in povzroči konformacijsko spremembo strukture potrebnega proteina, da se presnovi presnova na drugi strani membrane.
Vendar se vse te izoforme med seboj razlikujejo tako, da predstavljajo razlike v:
- Stopnjo afinitete, ki jo imajo za glukozo,
- Sposobnost izvajanja glukoze, galaktoze in aminokislin,
- Stopnjo, do katere jih zavirata Florizina in
- Lokacija tkiva.
[TOC]
Molekularni mehanizmi transporta glukoze
Glukoza je monosaharid šestih ogljikovih atomov, ki jih večina obstoječih tipov celic uporablja za pridobivanje energije s presnovnimi oksidacijskimi potmi.
Glede na svojo veliko velikost in v bistvu hidrofilno naravo ne more prečkati celičnih membran s prostim difuzijo. Zato je njegova mobilizacija citosola odvisna od prisotnosti transportnih beljakovin v teh membranah.
Do zdaj preučene glukozne prevozniki izvajajo prevoz tega presnovka s pasivnimi ali aktivnimi transportnimi mehanizmi. Pasivni transport se razlikuje od aktivnega prevoza, v katerem ne potrebuje oskrbe z energijo, saj se pojavi v prid koncentracijskemu gradientu.
Lahko vam služi: povrhnjica čebuleBeljakovine, vključene v pasivni transport glukoze, pripadajo družini razširjenih razširjenih prevoznikov, ki jih tako imenuje kratica v angleščini izraza "prevozniki glukoze". Medtem ko so tisti, ki izvajajo aktiven transport, imenovali SGLT z "natrijevim glukoznim transportnim beljakovinam".
Slednji dobijo potrebno prosto energijo za izvajanje transport glukoze glede na koncentracijski gradient natrijevega iona kotransport. Ugotovljenih je bilo vsaj 6 izoformov SGR in zdi se, da je njegova lokacija omejena na membrane epitelijskih celic.
Značilnosti SGLT
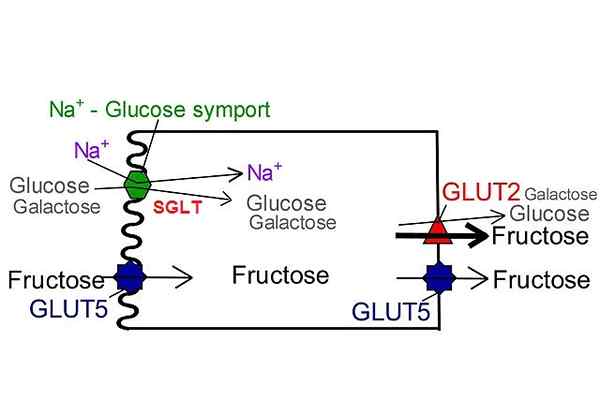
Kot je prikazano po imenu, transportni proteini natrijeve glukoze izvajajo simport tip natrija in glukoze ali natrija in galaktoze v celični citosol.
Prenos natrija v prid koncentracijskega gradienta je odgovoren za ustvarjanje nepogrešljive proste energije za mobilizacijo glukoze iz območja z nizko koncentracijo v visoko koncentracijo.
Ti transporterji so del skupine proti transportu in naklonjenosti natrija in vodika, katerih geni niso filogenetsko povezani.
Njegova lokacija je omejena na luminalno membrano epitelijskih celic tkiv, ki sodelujejo pri absorpciji hranil, kot sta tanek črevesje in proksimalni konturirani tubul.
Vam lahko služi: coxiella burnetiiVrste sglt
Do danes je bilo ugotovljenih šest prevoznikov SGLT (SGLT-1, SGLT-2, SGLT-3, SGLT-4, SGLT-5 in SGLT-6). Vsi kažejo razlike v preganjanju prevoza glukoze ali galaktoze, afiniteti, ki jo predstavljajo za te sladkorje in natrij, ter z dejstvom, da jih lahko zavira Florizina.
SGLT1 ima možnost prevoza galaktoze poleg glukoze z zelo podobno kinetiko, medtem ko SGLT2 samo prenaša glukozo.
GSLT struktura
Dolžina peptidnega zaporedja beljakovin SGLT se giblje od 596 do 681 aminokislinskih odpadkov. Medtem se odstotek homologije med sekvencami giblje med 50 in 84 % glede na SGLT-1, kar je največja stopnja razhajanja v sekvencah, ki ustrezajo zunajceličnemu domenskemu amino terminalu in terminalnem karboksilu.
Splošna struktura sovpada s strukturo integralnega proteina z multipasom, to je, da večkrat prečka membrano skozi transmembranske domene, bogate s hidrofobnimi aminokislinami.
Te transmembranske domene so skupno 14 in vsaka ima sekundarno strukturo α-helix. Segmenti 1, 2, 5, 7 in 9 so prostorsko organizirani v osrednjem položaju, kar povzroča nastanek hidrofilnih pore.
S hidrofilnimi pore tranzitira glukozo in tudi kateri koli drugi presnovki, za katere ima lahko transporter afiniteto. Preostali spiralni fragmenti so vzporedno organizirani, da se kanalu podelijo večjo strukturno stabilnost.
Sglt funkcije
Družinski člani SGLT ne kažejo edinstvene specifičnosti za glukozo. Nasprotno, lahko aktivno mobilizirajo skozi membrano celic ledvičnih tubul in črevesni epitelij široko paleto presnovkov, kot so aminokisline, ioni, glukoza in osmoliti.
Funkcija te vrste prevoznikov, ki je bila širše raziskana, je reabsorpcija glukoze, ki je prisotna v urinu.
Lahko vam služi: primerjalna embriologija: kaj je to, kakšne študije, zgodovina, teorijeTa reabsorpcijski postopek vključuje mobilizacijo ogljikovih hidratov iz ledvičnih tubulov skozi cevaste epitelijske celice do lumena peritubularnih kapilar. Ker je izoforma visoke zmogljivosti in afinitete do glukoze SGLT-2, ki je glavni davčni zavezanec.
Absorpcijska funkcija glukoze v črevesnem traktu se pripiše SGLT-1, transportniku, ki ima kljub nizki zmogljivosti visoko afiniteto do glukoze.
Tretji član te družine, SGLT3, je izražen v membranah skeletnih mišičnih celic in živčnega sistema, kjer se zdi, da ne deluje kot transporter glukoze, ampak kot senzor koncentracij tega sladkorja v zunajceličnem mediju.
Funkcije izoform SGLT4, SGLT5 in SGLT6 do zdaj še niso bile prepričane.
Reference
- Abramson J, Wright Em. Struktura in funkcija Na simporterjev z vklenjenimi ponovitvami. Curr Ace Struct Biol. 2009; 19: 425-432.
- Alvarado F, Crane RK. Študije o mehanizmu črevesne absorpcije sladkorjev. Vii. Fenilgllycozidni transport in njegov možni odnos do zaviranja forizina za transport Actice SUGS s majhnimi inštimi. Biochim Biophys Act. 1964; 93: 116-135.
- Charron FM, Blanchard MG, Lapointe JY. Intracelična hipertoničnost je odgovorna za vodni tok, povezan z Na_/glikozni kotransport. Biophys J. 2006; 90: 3546-3554.
- Chen XZ, Coady MJ, Lapointe JY. Hitro napetostne razpršitve v novi komponenti tokov perparAd-ity iz Na_-glukoze kotransporterja. Biophys J. Devetnajst devetdeset šest; 71: 2544-2552.
- Dyer J, Wood Is, Palejwala A, Ellis A, Shirazi-Beechey sp. Izražanje monosaharidnih transporterjev v črevesju diabetičnih ljudi. Am J Physiol najpogostejši jeter fiziol. 2002; 282: G241-G248.
- Sátk M, Marks J, Unwin RJ. Domnevna lokacija in funkcija družinskega člana SLC5 SGLT3. Exp Physiol. 2017; 102 (1): 5-13.
- Turk E, Wright Em. Motivi membranske topologije v družini SGG Cotransporter. J Memb Biol. 1997; 159: 1-20.
- Turk E, Kim O, Hils J, Whitelegge JP, Eskandari S, Lam JT, Kreman M, Zampighi G, Faull KF, Wright EM. Molekularna karakterizacija Vibrio parahaemolyticus VSGLT: model za natrijeve sladkorne kotransporte. J Biol Chem. 2000; 275: 25711-25716.
- Taroni C, Jones S, Thornton JM. Analiza in napovedovanje vezavnih mest za ogljikove hidrate. Eng protein. 2000; 13: 89–98.
- Wright Em, Loo DD, Hirayama BA. Biologija človeških natrijevih prevoznikov glukoze. Physiol Rev. 2011; 91 (2): 733-794.
- « Značilnosti zemeljske magnetosfere, struktura, plini
- Elementi magnetnega upada in zemeljsko magnetno polje »