

Strukture heksokinaze, funkcije, obrazci
- 2697
- 441
- Barry Ernser
The Heksokinaza (HK) je encim, ki katalizira prvo reakcijo glikolitične poti praktično v vseh živih organizmih, tako v prokariotih kot v evkariotih. Odgovoren je za prenos fosforilne skupine proti glukozi, ki proizvaja glukozo-6p, čeprav lahko fosforilira tudi druge hektozne sladkorje (6 ogljikovih atomov).
Ta encim je razvrščen v eno od dveh družin encimov glukoze -kinaze (encimi, ki prenašajo fosforilne skupine na substrate, kot je glukoza): družina heksokinaze (HK), katerih člani so razdeljeni v tri različne skupine, znane kot skupina HK, The HK Skupina A in skupina B.
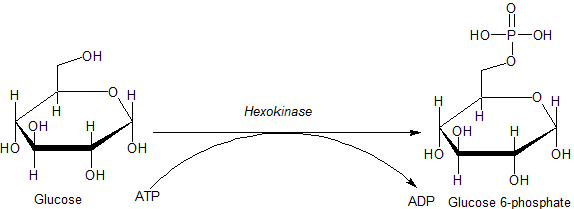
Reakcija, ki jo katalizira encim heksokinaze (vir: JMUN7616 [javna domena] prek Wikimedia Commons) Za encime, ki pripadajo družini HK, je značilna fosforilacija glukoze na račun ATP kot molekula donatorja fosforilne skupine, pripadniki pa se med seboj razlikujejo predvsem glede na njihovo molekulsko maso in specifičnost substrata.
Skupina HK pripada encimi evkariontskih organizmov (ATP: D-heksose 6-fosfotransferaze), medtem ko skupina A predstavljajo encimi gram-negativnih bakterij, cianobakterij, amitdomamatidne proste -Pozitivne bakterije in organizmi Creanarqueotas.
Encimi skupin A in B so znani tudi kot glikokinaze (GLCKS), saj so sposobni izključno glukozno fosforilacijo, zato se ti encimi imenujejo ATP: D-glukoza 6-fosfotransferaza.
Kot glikolitični encim ima heksohinaza velik presnovni pomen, saj brez nje ta pomembna pot ne bi bila mogoča in so celice, ki so zelo odvisne od uživanja ogljikovih hidratov na splošno.
[TOC]
Struktura
Kot bomo videli pozneje, pri sesalcih in drugih vretenčarjih (pa tudi v enoceličnih organizmih, kot so kvasovke), obstajajo različne vrste encimov heksokinaze. Pri sesalcih so bili opisani štirje: izoformi I, II, III in IV.
Vam lahko služi: železniške zlitine: značilnosti, vrste, primeriPrvi trije izoencimi imajo 100 kDa molekulsko maso, vendar ima izoencim IV 50 kDa. Ti izoencimi (zlasti I-III) kažejo veliko podobnost zaporedja med njima glede na njihov C- in N-terminalni konci ter druge člane družine heksokinaze.
N-terminalna domena teh encimov velja za "regulator" domeno, medtem ko katalitično aktivnost izvaja C-terminalna domena (HK II sesalcev ima aktivna mesta v obeh področjih).
N-terminalna domena je povezana s C-terminalnim domeno prek alfa propelerja, vsaka ima približno 50 kDa molekulsko maso in ima stičišče za glukozo.
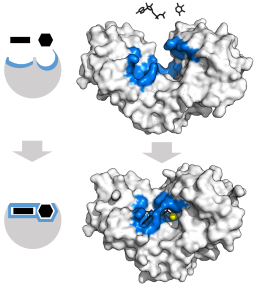 Inducirani model prilagoditve za encim heksokinaze (glede na dve podlagi: ATP in glukozo) (vir: Thomas Shafee [CC by 4.0 (https: // creativeCommons.Org/licence/by/4.0)] prek Wikimedia Commons)
Inducirani model prilagoditve za encim heksokinaze (glede na dve podlagi: ATP in glukozo) (vir: Thomas Shafee [CC by 4.0 (https: // creativeCommons.Org/licence/by/4.0)] prek Wikimedia Commons) Terciarna struktura teh encimov je v bistvu sestavljena iz β-nameščenih listov, pomešanih z alfa propelerji, katerih delež se razlikuje glede na encim in zadevne vrste; Stranjo sindikata za ATP, drugi heksokinazni substrat, je običajno sestavljeno iz petih β listov in dveh alfa propelerjev.
Funkcije
Heksokinaza ima transcendentalno funkcijo v presnovi ogljikovih hidratov večine živih bitij, saj katalizira prvi korak glikolitične poti, ki posreduje fosforilacijo glukoze v notranjosti.
Ta prvi korak glikolize, ki je sestavljen iz prenosa fosforilne skupine iz ATP (darovalca) v glukozo, ki daje 6-fosfat in ADP glukozo, je prvi od obeh korakov naložbe v energijo v obliki ATP.
Poleg tega je reakcija, ki jo katalizira heksokinaza, prehod "aktivacije" glukoze za nadaljnjo obdelavo in predstavlja korak "zavezanosti".
Vam lahko služi: heterogene mešaniceProdukt reakcije, ki jo katalizira heksokinaza, to je 6-fosfatna glukoza, je točka veje, saj je prvi substrat, ki se uporablja na poti pentoznega fosfata in v sintezi glikogena pri mnogih živalih (in o škrob na rastlinah).
V rastlinah
Funkcija heksokinaze v rastlinah se ne razlikuje zelo od funkcije živali ali mikroorganizmov, vendar v zgornjih rastlinah ta encim deluje tudi kot "senzor" koncentracije sladkorja.
Pomen te funkcije v teh organizmih je povezan s sodelovanjem sladkorjev kot regulacijskih dejavnikov pri izražanju genov, ki sodelujejo v različnih presnovnih procesih, kot so:
- Fotosinteza
- Cikel glioksilata
- Dihanje
- Razgradnja ali sinteza škroba in saharoze
- Presnova dušika
- Obramba pred patogeni
- Regulacija celičnega cikla
- Zdravilni odziv
- Pigmentacija
- Med drugim.
Ta funkcija heksokinaze kot "senzorja" količine znotrajcelične glukoze je bila opisana tudi za kvasovke in sesalce.
Oblike
V naravi obstajajo različne oblike heksokinaz, kar je v bistvu odvisno od vrste, ki se upoštevajo.
Na primer pri človeških in drugih živalih je bilo dokazano obstoj štirih različnih izoform encima heksokinaze v citosolnem oddelku, ki so ga označevali z rimskimi številkami I, II, III in IV.
Isoencimi, II in III sta 100 kDa molekulska masa, zavirajo njihov reakcijski produk. Vendar imajo ti encimi malo specifičnosti substrata, saj lahko fosforilirajo druge heksoze, kot sta fruktoza in roka.
Vam lahko služi: kisline: značilnosti in primeriIsoencim IV, znan tudi kot glikokinaza (GLCK), ima le 50 kDa molekulsko maso in ima kljub temu, da je malo povezan (visoke vrednosti km) Trije izoencimi.
Glukokinaza (izoencim IV heksokinaze številnih sesalcev) najdemo predvsem v jetrih in pomaga temu organu pri "prilagoditvi" hitrosti porabe glukoze kot odziv na spremembe tega substrata v obtočni krvi.
Trije geni, ki kodirajo za heksokinaze I, II in III pri živalih, se zdijo, da III prebivajo le na koncu C-terminala.
Reference
- Aronoff, s. L., Berkowitz, k., Shreiner, b., & Hoče, l. (2004). Metabolizem in regulacija glukoze: onstran insulina in glukagona. Spekter diabetes, 17(3), 183-190.
- Harrington, g. N., & Bush, D. R. (2003). Bifuncialna vloga heksokinaze pri presnovi in glukozni signalizaciji. Rastlinska celica, petnajst(11), 2493-2496.
- Jang, j. C., León, str., Zhou, l., & Sheen, J. (1997). Heksokinaza glede senzorja sladkorja v višjih rastlinah. Rastlinska celica, 9(1), 5-19.
- Kawai, s., Mukai, t., Mori, s., Mikami, b., & Murata, k. (2005). Hipoteza: strukture, evolucija in prednik glukoznih kinaz v družini heksokinaze. Časopis za bioznanost in bioinženiring, 99(4), 320–330.
- Mathews, k. In. (1998). Van Holde. Biokemija.
- Wilson, J. In. (2003). Isocimi heksokinaze sesalcev: struktura, podcelična lokalizacija in presnovna funkcija. Časopis za eksperimentalno biologijo, 206(12), 2049–2057.
- « 13 sokov za zdravljenje gastritisa (naravni in učinkoviti)
- Vzroki in primeri divergentne evolucije »