

Funkcije, vrste in primeri restrikcijskih encimov
- 3215
- 94
- Stuart Armstrong
The Restrikcijski encimi So endonukleaze, ki jih nekateri loki in bakterije uporabljajo za zaviranje ali "omejevanje" širjenja virusov v notranjosti. Posebej so pogosti v bakterijah in so del njihovega tujega obrambnega sistema DNK, znan kot sistem omejitve/spreminjanja.
Ti encimi katalizirajo dvojno pasovno DNK na določenih mestih, ponovljivo in brez uporabe dodatne energije. Večina zahteva prisotnost kofaktorjev, kot so magnezij ali drugi dvovalentni kationi, čeprav nekateri potrebujejo tudi ATP ali S-adenozil metionin.
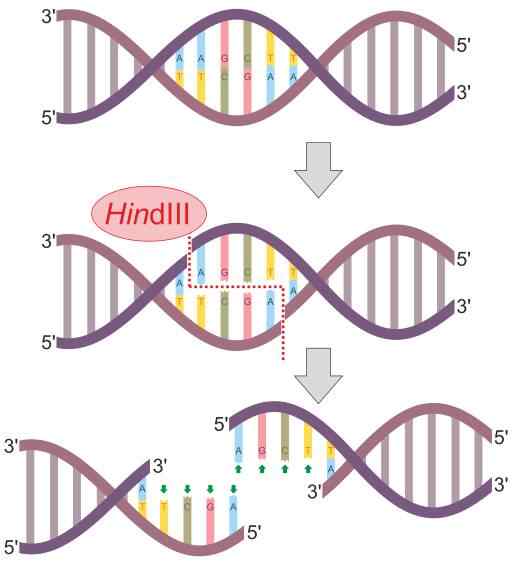
Shema reakcijske reakcijske reakcije hinduii (vir: Helixitta [CC BY-SA 4.0 (https: // creativeCommons.Org/licence/by-sa/4.0)] prek Wikimedia Commons) Restrikcijske endonukleaze so leta 1978 odkrili Daniel Nathans, Arber Werner in Hamilton Smith, ki so v medicini prejeli Nobelovo nagrado za svoje odkritje. Njegovo ime na splošno izhaja iz organizma, kjer so jih prvič opazili.
Takšni encimi se pogosto uporabljajo pri razvoju metod kloniranja DNK in drugih molekularnih biologij in strategij genetskega inženirstva. Njegove značilnosti prepoznavanja specifičnih zaporedij in rezalne zmogljivosti zaporedja, ki so blizu mesti prepoznavanja.
Fragmenti, ki jih ustvarijo restrikcijski encimi, ki so delovali na določeni molekuli DNK.
Nekateri restrikcijski encimi imajo lahko enako mesto prepoznavanja v DNK, vendar ga ne režejo nujno na enak način. Tako obstajajo encimi, zaradi katerih se reže zapuščajo Romos in encimi, ki režejo kohezivne skrajnosti, ki imajo različne aplikacije v molekularni biologiji.
Trenutno je na voljo na stotine različnih restrikcijskih encimov, ki jih ponujajo različne poslovne hiše; Ti encimi delujejo kot "prilagojene" molekularne škarje za različne namene.
[TOC]
Funkcije
Restrikcijski encimi izpolnjujejo nasprotno funkcijo polimeraz, saj so hidrolizirani ali razbijejo ester vez znotraj fosfodiésterjeve vezi med sosednjimi nukleotidi v nukleotidni verigi.
V molekularni biologiji in genetskem inženiringu se široko uporabljajo za gradnjo izražanja in kloniranja, pa tudi za identifikacijo specifičnih zaporedij. Uporabni so tudi za gradnjo rekombinantnih genomov in imajo velik biotehnološki potencial.
Nedavni napredek v genski terapiji trenutno uporablja restrikcijske encime za uvedbo genov, določenih v vektorjih, ki so vozila za prevoz takšnih genov v žive celice, in da se verjetno lahko vstavijo v celični genom, da bi izvedli trajne spremembe.
Mehanizem delovanja
Restrikcijski encimi lahko katalizirajo rez DNA z dvojnim pasom, čeprav so nekateri sposobni prepoznati preproste sekvence DNK in celo RNA. Rez se pojavi po prepoznavanju zaporedij.
Vam lahko služi: cikel glioksilata: značilnosti, reakcije, regulacija, funkcijeMehanizem delovanja je sestavljen iz hidrolize fosfodiésterjeve povezave med fosfatno skupino in deoksiribozo v okostju vsake DNK DNK. Številni encimi so sposobni rezati na isto mesto, kot ga prepoznajo, drugi pa so se prerezali med 5 in 9 osnovnih parov pred ali po istem.
Običajno ti encimi režejo na 5 'koncu fosfatne skupine, kar povzroča fragmente DNK s 5' fosforilnim koncem in terminalom hidroksilnega konca 3 '.
Ker beljakovine ne pridejo v neposreden stik s mestom za prepoznavanje DNK, jih je treba premeščati zaporedni časi, dokler ne dosežemo določenega mesta, morda z "drsnimi" mehanizmi na pramentu DNK.
Med encimskim rezom je fosfodiésterska povezava vsakega od pramenov DNK nameščena na enem od aktivnih mest restrikcijskih encimov. Ko encim zapusti mesto prepoznavanja in rezanja, to stori prek ne -specifičnih prehodnih asociacij.
Fantje
Trenutno je znanih pet vrst restrikcijskih encimov. Nato kratek opis vsakega:
Restrikcijske encime tipa I
Ti encimi so veliki pentamerni proteini s tremi podenotami, ena omejitev, metilacija in drugo za prepoznavanje sekvenc DNK. Te endonukleaze so večnamenski proteini, ki so sposobni katalizirati reakcije na omejevanje in spreminjanje, imajo ATPASA in tudi aktivnost topoizomera DNA.
Encimi te vrste so bili prve endonukleaze, ki so jih odkrili, očistili so jih prvič v šestdesetih letih prejšnjega stoletja in od takrat so jih preučevali z veliko globino.
Encimi tipa I se ne uporabljajo široko kot biotehnološko orodje, saj je mesto rezanja lahko na spremenljivi razdalji do 1.000 osnovnih parov glede mesta prepoznavanja, zaradi česar so nezanesljivi v smislu eksperimentalne obnovljivosti.
Restrikcijski encimi tipa II
So encimi, sestavljeni iz homodimerov ali tetramerov, ki DNK režejo na mesta, določena med 4 in 8 bp dolžine. Ta mesta za rezanje so običajno palindromska, torej prepoznavajo sekvence, ki se berejo na enak način v obe smeri.
Številni restrikcijski encimi tipa II v bakterijah so prerezali DNK, ko prepoznajo svoj tuji značaj, saj nima značilnih sprememb, ki bi jih moral imeti lastni DNK.
To so preprostejši restrikcijski encimi, saj ne potrebujejo nobenega kofaktorja, razen magnezija (Mg+), da prepoznajo in režejo sekvence DNK.
Natančnost restrikcijskih encimov tipa II pri prepoznavanju in rezanju preprostih zaporedij v DNK v natančnih položajih je ena najbolj uporabljenih in nepogrešljivih v večini vej molekularne biologije.
Vam lahko služi: vzajemnost: značilnosti, vrste, primeriV skupini restrikcijskih encimov tipa II je več podrazred, razvrščenih v skladu z določenimi lastnostmi, ki so edinstvene za vsakega. Klasifikacija teh encimov se izvede z dodajanjem črk abecede, od A do Z po imenu encima.
Nekateri najbolj znani podrazred za njihovo uporabnost so:
Podrazred Iia
So različna podenota dímeros. Prepoznajo asimetrične sekvence in se uporabljajo kot idealni predhodniki za ustvarjanje rezalnih encimov.
Podrazred IIB
Sestavljeni so iz še enega dimerjev in razrežejo DNK na obeh straneh zaporedja prepoznavanja. Obe prameni DNK so v intervalu baznih parov pozneje izrezali s mesta prepoznavanja.
IIC podrazred
Encimi te vrste so polipeptidi z delitvijo in spreminjajočimi funkcijami pramenov DNK. Ti encimi asimetrično razrežejo oba pramena.
Podrazred iie
Encimi tega podrazreda se najbolj uporabljajo v genetskem inženiringu. Imajo katalitično mesto in na splošno potrebujejo alestrovski efektor. Ti encimi morajo za učinkovito rezanje komunicirati z dvema kopijama njihovega zaporedja prepoznavanja. Znotraj tega podrazreda so encimi Ecorii in Ecori.
Restrikcijski encimi tipa III
Endonukleaze Type III so sestavljene samo iz dveh podenot, ena je odgovorna za prepoznavanje in spreminjanje DNK, druga.
Ti encimi za njihovo delovanje potrebujejo dva kofaktorja: ATP in magnezij. Tovrstni restrikcijski encimi imajo dve asimetrični mesti prepoznavanja, DNK premestijo na ATP -odvisen način in jo razrežete med 20 in 30 bp v bližini mesta prepoznavanja.
Restrikcijski encimi tipa IV
Encime tipa IV je enostavno prepoznati, saj režejo DNK z metilacijskimi oznakami, sestavljajo jih več različnih podenot, ki so odgovorni za prepoznavanje in rezanje zaporedja DNK. Ti encimi uporabljajo kofaktorje GTP in dvovalentni magnezij.
Specifična mesta za rezanje vključujejo nukleotidne verige z ostanki citozina z metiliranimi ali hidroksimesiji v enem ali obeh pramenih nukleinskih kislin.
Restrikcijski encimi tipa V
Ta klasifikacijska skupina Crisper-Cas tipa encimi, ki prepoznavajo in režejo specifične sekvence DNK invazivnih organizmov. CAS Encimi uporabljajo RNA pramen sintetizirani vodnik za Crisper za prepoznavanje in napad na invazivne organizme.
Encimi, razvrščeni kot tipa V, so polipeptidi, strukturirani po encimi tipa I, II in II. Lahko razrežejo odseke DNK skoraj vsakega organizma in z velikim razponom dolžine. Njegova prilagodljivost in enostavnost zaposlitve naredijo ti encimi eno najbolj uporabljenih orodij v genetskem inženiringu, ki je trenutno skupaj z encimi tipa II.
Vam lahko služi: propionibacteriumPrimeri
Restrikcijski encimi so bili uporabljeni za odkrivanje polimorfizmov DNA, zlasti v populacijskih raziskavah genetike in evolucijskih študijah z uporabo mitohondrijske DNK, da bi pridobili informacije o stopnji substitucije nukleotida.
Trenutno imajo vektorji, ki se uporabljajo za transformacijo bakterij z različnimi namesti.
Med temi encimi so najbolj priljubljeni Ecori, II, III, IV in V, pridobljeni in opisani prvič od In. coli; Hinduii, od H. influenzae in bamhi iz B. Amiloliquefaciens.
Reference
- Bikle, t. Do., & Kruger, D. H. (1993). Biologija omejitve DNK. Mikrobiološki pregledi, 57(2), 434–450.
- Boyaval, str., Moineau, s., Romero, d. Do., & Horvath, str. (2007). CRISPR zagotavlja pridobitev proti virusom v prokariotih. Znanost, 315(Marec), 1709-1713.
- Borgesell, d. (2002). Molekularna perspektiva: omejevalne endonukleaze. Osnove matičnih celic medicine raka, dvajset, 190-191.
- Halford, s. In. (2001). Skok, skakanje in zanko z restrikcijskimi encimi. Transakcije z biokemično družbo, 29, 363-373.
- Jeltsch, a. (2003). Vzdrževanje identitete vrst in nadzor specifikacije bakterij: nova funkcija za sisteme za omejevanje/spreminjanje? Gen, 317, 13-16.
- Krebs, j., Goldstein, e., & Kilpatrick, s. (2018). Lewinovi geni xii (12 izd.). Burlington, Massachusetts: Jones & Bartlett Learning.
- Li, in., Kruh, s., Zhang in., Ren, m., Feng, m., Peng, n.,... ona, Q. (2015). Izkoriščanje sistemov tipa I in tipa III CRISPR-CAS za urejanje genoma. Raziskave nukleinskih kislin, 1-12.
- Loenen, w. Do. M., Dryden, d. T. F., Raleight, e. Do., & Wilson, g. G. (2013). Restrikcijski encimi tipa I in njihovi sorodniki. Raziskave nukleinskih kislin, 1-25.
- Nathans, d., & Smith, h. Tudi. (1975). Restrikcijske endonukleaze pri analizi in prestrukturiranju molekul DNK. Annu. Rev. Biochem., 273-293.
- Nei, m., & Tajima, f. (1981). DNK polimorfizem, ki ga je mogoče zaznati z restrikcijskimi endonukleatami. Genetika, 145-163.
- Pingoud, a., Fuxreiter, m., Pingoud, v., & Wende, w. (2005). Celične in molekularne življenjske vede Endonukleaze tipa II: struktura in mehanizem. CMLS celične in molekularne znanosti o življenju, 62, 685-707.
- Roberts, r. (2005). Kako so restrikcijski encimi postali delovni konji molekularne biologije. PNAS, 102(17), 5905-5908.
- Roberts, r. J., & Murray, k. (1976). Omejene endonukleaze. Kritični pregledi v biokemiji, (November), 123-164.
- Stoddard, b. L. (2005). Homing endonukleazna struktura in funkcija. Četrtletne ocene biofizike, 1-47.
- Tock, m. R., & Dryden, D. T. F. (2005). Biologija omejitve in protiportrecije. Trenutno mnenje v mikrobiologiji, 8, 466-472. https: // doi.org/10.1016/j.Moj b.2005.06.003
- Wilson, g. G., & Murray, n. In. (1991). Sistemi za omejevanje in spreminjanje. Annu. Rev. Genet., 25, 585-627.
- Wu, z., & Mou, k. (2016). Genomski vpogled v virulenco Campylobacter Jejuni in populacijsko genetiko. Infec. Dis. Prevajalec. Med., 2(3), 109-19.
- Juan, r. (1981). Struktura in mehanizem večnamenskih omejitev endonukleat. Annu. Rev. Biochem., petdeset, 285-315.
- « Generacija značilnosti iz leta 1914, avtorji in dela
- Medsebojno izključujoče lastnosti dogodkov in primeri »