

Uracilna struktura, funkcije, lastnosti, sinteza
- 3926
- 271
- Adrian Legros
On Uracil Gre za pirimidinsko dušikovo bazo, ki jo najdemo v ribonukleinski kislini (RNA). To je ena od značilnosti, ki razlikuje RNA od deoksiribonukleinske kisline (DNK), saj ima slednja timina namesto Uracila. Obe snovi, Uracil in Timina, se razlikujeta le, da ima drugo metilno skupino.
Z evolucijskega vidika je bilo predlagano, da je RNA prva molekula, ki je shranjevala genetske informacije in delovala kot katalizator v celicah, pred DNK in encimi. Zaradi tega se misli, da je imel Uracil ključno vlogo pri razvoju življenja.
Vir: Kemikungen [javna domena] V živih bitjih uracila ne najdemo prosto, ampak pogosto monofosfatni nukleotidi (UMP), difosfat (UDP) in Trphosfat (UTP). Ti uracilni nukleotidi imajo različne funkcije, kot sta biosinteza RNA in glikogen, izomerna medkonverzija sladkorjev in regulacija sintaze sintaze.
[TOC]
Struktura in lastnosti
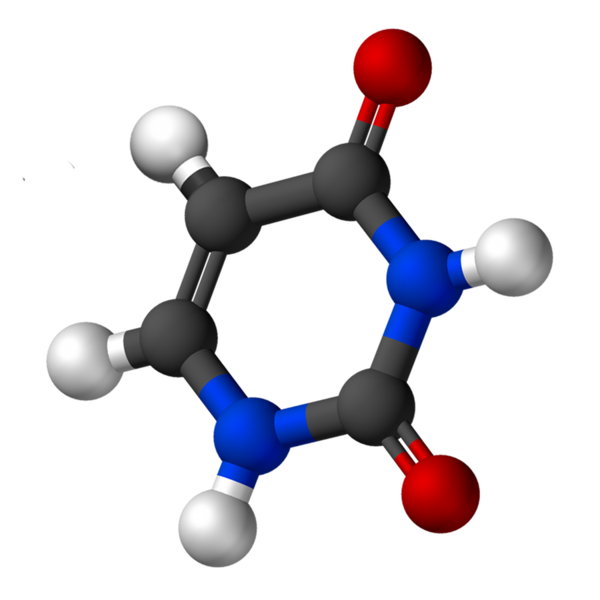
Uracil, imenovan 2.4-dioksipiridin, ima empirično formulo c4H4N2Tudi2, katere molekulska teža je 112,09 g/mol in je očiščena kot beli prah.
Struktura uridina je heterociklični obroč s štirimi ogljikovimi atomi in dvema dušikoma, z dvojnimi nadomestnimi vezmi. Je planarno.
Ima 50 mg/ml topnosti, 25 ° C, v 1M natrijevem hidroksidu in PKA med 7.9 in 8.2. Valovna dolžina, kjer se pojavi njegova največja absorbanca (ʎMax) Je med 258 in 260 nm.
Biosinteza
Obstaja skupna pot za biosintezo pirimidinskih nukleotidov (uracil in citokin). Prvi korak je biosinteza karbamoil fosfata iz CO2 in nh4+, ki ga katalizira fosfat s karbamoil sintetizira.
Pirimidin je zgrajen iz ogljikovega fosfata in aspartata. Obe snovi reagirajo in tvorita n-carbamoilaspartato, reakcija, ki jo katalizira transcabamoilasa aspartat (ATCASA). Zaprtje pirimidinskega obroča lahko dehidriramo z dihidrootazo in ustvari l-dihidrotat.
L-dihidrotoat se oksidira in spremeni v orotat; Acceptor elektronov je NAD+. Je reakcija, ki jo katalizira dihidroorotat dehidrogenaza. Naslednji korak je sestavljen iz prenosa fosforozilne skupine, iz fosforibozil pirofosfata (PRPP), do orotata. Oblika GradiDilato (WPO) in anorganski piroffosfat (PPI), ki jo katalizira fosfoshozil orotat transferaza.
Zadnji korak je sestavljen iz dekarboksilacije pirimidinskega obroča skupine (WPO). Uridilaato oblika (uridin-5'-monophosfat, UMP), ki jo katalizira dekarboksilaza.
Nato se s sodelovanjem kinaze fosfatna skupina prenese iz ATP v UMP in tvori UDP (Uridin-5'-difosfat). Slednje se ponovi in tvori UTP (Uridin-5'-Triffosfat).
Vam lahko služi: veje biologije in kaj študirajoUredba o biosintezi
V bakterijah se regulacija biosinteze pirimidina pojavi z negativnimi povratnimi informacijami, na ravni transcabamoilasasasa (ATCASA).
Ta encim zavira CTP (Citidin-5'-Trophosfat), ki je končni produkt biosinteze poti pirimidinov. ATCAS.
Pri živalih se regulacija biosinteze pirimidina zgodi z negativnimi povratnimi informacijami, na ravni dveh encimov: 1) fosfat karbamoil sintaze II, ki ga zavirata UTP in aktivirata ATP in PRPP; in 2) OMP diskorboksilaza, ki jo zavira produkt reakcije, ki jo sama katalizira, UMP. Hitrost biosinteze OMP se razlikuje glede na razpoložljivost PRPP.
Delovanje v biosintezi RNA
Uracil je prisoten v vseh vrstah RNA, kot so messenger RNA (MNA), prenosna RNA (ARNT) in ribosomalna RNA (RNA). Biosinteza teh molekul se zgodi s postopkom, imenovanim transkripcija.
Med prepisom se podatki, ki jih vsebuje DNK, kopirajo v RNA prek RNA Pon polimeraze. Obrni postopek, v katerem se informacije, ki jih vsebuje RNA, kopirajo v DNK, se zgodi v nekaterih virusih in rastlinah skozi obratno transkriptazo.
Biosinteza RNA potrebuje triposfatne nukleozide (NTP), in sicer: triposfatni citidin (CTP), adenin triposfat adenin (ATP) in triposfat gvanin (GTP) (GTP) (CTP) (GTP). Reakcija je:
(RNA)n odpadki + NTP -> (RNA)N+1 odpadki + ppi
Hidroliza anorganskega pirofosfata (PPI) zagotavlja energijo za biosintezo RNA.
Delovanje v biosintezi sladkorja
Sladkorni estri so zelo pogosti v živih organizmih. Nekateri od teh estrov so difosfe nukleozidnih estrov, kot je UDP-Sugar, ki so v celicah zelo obilne. UDP-Sugar vključuje biosintezo disaharidov, oligosaharidov in polisaharidov.
V rastlinah se biosinteza saharoze zgodi na dva načina: glavni in sekundarni.
Glavna pot je sestavljena iz prenosa D-glukoze iz UDP-D-glukoze v D-Fruceazo, da tvori saharozo in UDP. Sekundarna cesta vključuje dva koraka: začne se z UDP-D-glukozo in fruktozo-6-fosfatom in vrhunec s tvorbo saharoze in fosfata.
V mlečnih žlezah se biosinteza laktoze zgodi iz UDP-D-galaktoze in glukoze.
Vam lahko služi: veje biokemijeV rastlinah se biosinteza celuloze izvaja z neprekinjeno kondenzacijo beta-D-glikozilnih odpadkov, od UDP-glukoze do ne-zmanjševanja konca rastočega poliglukozne verige. Podobno je za biosintezo amiloze in amilopektina potrebna UDP-glukoza kot substrat darovalca glukoze do rastoče verige.
Pri živalih se za biosintezo glikogena uporabljata tako UDP-glukoza kot ADP-glukoza. Podobno za biosintezo hondroitina sulfata zahteva UDP-ksilozo, UDP-galaktozo in UDP-glukuronato.
Funkcija v izomerni medsebojni pretvorbi sladkorjev
Pretvorba galaktoze v posrednik glikolize se zgodi skozi cesto Leloir. Eden od korakov te poti katalizira encim UDP-galaktoze-4-epimeraze, ki olajša medsebojno pretvorbo UDP-galaktoze do UDP-glukoze.
Delovanje v biosintezi glikoproteinov
Med biosintezo glikoproteinov beljakovine gredo skozi cis, srednje in trans vreče Golgijevega aparata.
Vsaka od teh vrečk ima nabor encimov, ki obdelujejo glikoproteine. Sladkorni monomeri, kot sta glukoza in galaktoza.
Nukleotide-heksose se z antiportom prevaža v rezervoarje Golgi. UDP-galaktoza (UDP-GAL) in UDP-N-acetilgalaktosamina (UDP-GalNAC) vstopata v rezervoarje iz citosola s pomočjo izmenjave.
V rezervoarju Golgi fosfataza hidrolizira fosfatno skupino oblike UDP in UMP ter PI. UDP izvira iz reakcij, ki jih katalizirata galaktosiltransferaza in N-acetilgalaktosamiltransferaza. UMP, ki ga tvori fosfataza, služi za izmenjavo nukleotidov-heksoze.
Funkcija pri regulaciji sintaze
Mehanizem za uravnavanje glutaminske sintaze je kovalentna sprememba, ki je sestavljena iz adenilacije, ki neaktivna, in bok, ki jo aktivira. Ta kovalentna sprememba je reverzibilna in katalizira adeniltransferaza.
Aktivnost adeniltransferaze modulira Združenje proteina PII, ki ga uravnava kovalentna sprememba, uridinilacija.
Uridililacija in odmik izvede uridiltransferaza. V tem encimu je uridilacijska aktivnost posledica glutamina in fosfata, aktivira pa jo združitev alfa-zotoglutarata in ATP na PII.
Funkcija v izdaji RNA
Nekateri RNM so urejeni pred prevodom. V nekaterih evkariontskih organizmih, kot so Trypanosoma brucei, Obstaja izdaja RNA transkripta gena podenote II citokroma oksidaze. To se zgodi z vstavljanjem uracilnih odpadkov, reakcije, ki jo katalizira končna uridiltransferaza.
Vam lahko služi: curl: sestava, deli, pomenVodilna RNA, ki dopolnjuje urejen izdelek, deluje kot zmerna za postopek urejanja. Osnovni pari, oblikovani med začetnim prepisom in vodilnim RNA, pomeni osnovni pari g = u, ki niso Watson-record in so pogosti v RNA.
UDP-glukozna biosinteza
V fizioloških pogojih je biosinteza glikogena iz glukoze-1-fosfata termodinamično nemogoča (pozitiven ΔG). Zaradi tega se pred biosintezo zgodi aktiviranje glukoze-1-fosfata (G1P). Ta G1P in UTP združuje reakcijo, da tvori glukozni difosfat uridin (UDP-glukoza ali UDPG).
Reakcijo katalizira pirofosforilaza UDP-glukoze in je naslednja:
G1P + UTP -> UDP -Glucosa + 2pi.
Gibbs Prosti energiji v tem koraku je velika in negativna (-33,5 kJ/mol). Med reakcijo kisika G1P napade alforo alfa fosfor atom UDP-glukoze in anorganskega pirofosfata (PPI). Nato PPI hidrolizirajo anorganski pirofosfati, katerih energija hidrolize je tista, ki poganja splošno reakcijo.
UDP-glukoza je "visoko energijska" snov. Omogoča oblikovanje glikoznih vezi med ostankom glukoze in naraščajočo verigo polisaharida. To isto energijsko načelo je uporabno za reakcije, kjer sodeluje UDP-Sugar, na primer biosinteza disaharidov, oligosaharidov in glikoproteinov.
Uracil DNA Glycosilasa
Obstajajo lezije DNK, ki se pojavljajo spontano. Ena od teh lezij je smontane. V tem primeru popravilo poteka zaradi spremenjene baze DNK s pomočjo encima, imenovanega Uracil DNA glikozilasa.
Encim Uracil DNA glikosilasa odpravi poškodovan citokin (Uracil), kar proizvaja ostanek deoksiriboze, ki nima dušikove baze, imenovane AP mesto (apurinsko-apirimidinsko mesto).
Nato encim Endonuclease AP naredi rezanje v fosfodiesterskem okostju mesta AP, kar odpravi ostanke sladkorja-fosfata. DNK polimeraza obnavljam poškodovani pramen.
Reference
- Bohinski, r. 1991. Biokemija. Addison-Wesley Iberoamericana, Wilmington, Delaware.
- Devlin, t.M. 2000. Biokemija. Uredništvo Reverté, Barcelona.
- Lodish, h., Berk, a., Zipurski, s.L., Matsudaria, str., Baltimore, d., Darnell, J. 2003. Celična in molekularna biologija. Uredništvo Medica Panamericana, Buenos Aires, Bogota, Caracas, Madrid, Mehika, Sāo Paulo.
- Nelson, d. L., Cox, m. M. 2008. Lehning-principles biokemije. W.H. Freeman, New York.
- Voet, d. in voet, j. 2004. Biokemija. John Wiley in sinovi, ZDA.