

Sinteza lipidnih vrst in njihovih glavnih mehanizmov
- 2548
- 686
- Ms. Pablo Lebsack
The Sinteza lipidov Sestavljen je iz niza encimskih reakcij, s katerimi se ogljikovodiki kratkih verig kondenzirajo, da tvorijo daljše verižne molekule, ki lahko pozneje utrpijo različne kemijske spremembe.
Lipidi so zelo raznolike vrste biomolekul, ki jih sintetizirajo vse žive celice in so specializirane za več bistvenih funkcij za vzdrževanje življenjske dobe celic.
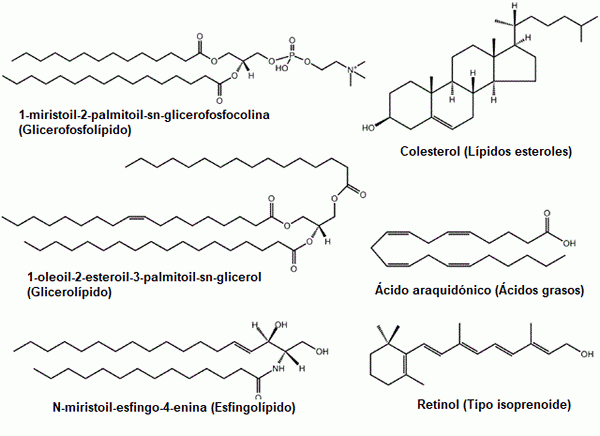 Nekaj primerov običajnih lipidov: glicerofosfolipidi, sterole, glicerolipidi, maščobne kisline, sfingolipidi in prenoli (vir: prvotni nalagalnik je bil LMAPS pri angleški Wikipediji. / Gfdl 1.2 (http: // www.GNU.Org/licence/stare licence/fdl-1.2.html) prek Commons, prilagodil Raquel Parada)
Nekaj primerov običajnih lipidov: glicerofosfolipidi, sterole, glicerolipidi, maščobne kisline, sfingolipidi in prenoli (vir: prvotni nalagalnik je bil LMAPS pri angleški Wikipediji. / Gfdl 1.2 (http: // www.GNU.Org/licence/stare licence/fdl-1.2.html) prek Commons, prilagodil Raquel Parada) Lipidi so glavne sestavine bioloških membran, dejstvo, zaradi katerega so temeljne molekule za obstoj celic kot izolirane entitete njihove okolice.
Nekateri lipidi imajo tudi specializirane funkcije, kot so pigmenti, kofaktorji, transporterji, detergenti, hormoni, intra-ekstracelični glasniki, kovalentna sidra za membranske beljakovine itd. Zato je sposobnost sinteze različnih vrst lipidov ključna za preživetje vseh živih organizmov.
Ta velika skupina spojin je tradicionalno razvrščena v več kategorij ali podskupin: maščobne kisline (nasičene in nenasičene), gliceridni kompleksni lipidi (lipoproteini).
[TOC]
Vrste lipidov in njihovi glavni mehanizmi sinteze
Vse reakcijske sekvence lipidnih biosinteznih poti so endogonske in reduktivne. Z drugimi besedami, vsi uporabljajo ATP kot vir energije in zmanjšanega transporterja elektronov, kot je NADPH.
Nato bodo opisane glavne reakcije biosintetskih poti glavnih vrst lipidov, torej maščobnih in eikosanoidnih kislin, triacilglicerolov in fosfolipidov in sterolov (holesterol) (holesterol).
- Sinteza maščobnih kislin
Maščobne kisline so izredno pomembne molekule z lipidnega vidika, saj so del najpomembnejših lipidov v celicah. Njegova sinteza, v nasprotju s tem, kar so mnogi znanstveniki mislili med prvimi študijami, ni sestavljena iz obratne poti njihove β-oksidacije.
Pravzaprav se ta presnovna pot pojavlja v različnih celičnih oddelkih in zahteva sodelovanje posrednika treh ogljikovih atomov, znanih kot malonil-CoA, kar pri oksidaciji ni potrebno.
 Malonil-CoA. Neuroteker / pub domena
Malonil-CoA. Neuroteker / pub domena Poleg tega je tesno povezan s skupinami sulfhidrilnih beljakovin, znanih kot transporterji acilnih skupin (ACP, angleščina Acil nosilni proteini).
Na splošno je sinteza maščobnih kislin, zlasti dolge verige, zaporedni postopek, kjer se v vsaki "vrnitvi ponovijo štiri korake, med vsakim donosom pa je nasičena kislinska skupina, ki je podlaga za naslednjo, ki je naslednja, ki je naslednja, ki je naslednja, ki je naslednja, ki je naslednja pomeni še eno kondenzacijo z novo molekulo malonil-CoA.
V vsakem zavoju ali reakcijskem ciklu se veriga maščobnih kislin podaljša dva ogljika, dokler ne doseže dolžina 16 atomov (palmitat), po katerem cikel odide.
Formacija alonila-CoA
Ta posrednik treh ogljikovih atomov je nepovratno oblikovan iz acetil-CoA, zahvaljujoč delovanju encima acetil-CoA karboksilaze, ki ima biotinsko skupino prostate, ki je kovalentno povezana z encimom in ki v tej katalizi v dveh korakih sodeluje v tej katalizi.
V tej reakciji se karboksilna skupina, ki izhaja iz molekule bikarbonata (HCO3-), prenese v biotin v obliki, odvisna od ATP, kjer biotinilna skupina izpolnjuje funkcijo "začasnega transporta" molekule, medtem ko se prenaša na acetil-CoA, proizvajajo malonil-CoA.
Vam lahko služi: test oksidaze: temelj, postopek in uporabeV zaporedju sinteze maščobnih kislin je uporabljeno sredstvo, ki se uporablja.
V vretenčarjih je kompleks maščobnih kislin sintaze del ene same velike polipeptidne verige, v kateri je zastopanih 7 encimskih aktivnosti, značilnih za sintezno pot, pa tudi hidrolitna aktivnost, potrebna za sproščanje posrednikov na koncu sinteze.
 Sintasa Encimska struktura maščobnih kislin (vir: Boehringer Ingelheim/CC BY-SA (https: // createCommons.Org/licence/by-sa/4.0) prek Wikimedia Commons)
Sintasa Encimska struktura maščobnih kislin (vir: Boehringer Ingelheim/CC BY-SA (https: // createCommons.Org/licence/by-sa/4.0) prek Wikimedia Commons) 7 encimskih aktivnosti tega kompleksa je: acilo skupinski transportni protein (ACP), acetil-CoA-ACP Transacetilas cetoacil-ACP reduktaza (KR), β-hidroksiacil-ACP dehidratasa (HD) in ANGER-ACP reduktaza (ER).
Preden se lahko pojavijo kondenzacijske reakcije, da sestavijo verigo maščobnih kislin, sta dve tiol skupini v encimskem kompleksu "naložene" z acilnimi skupinami: najprej se acetil -CoA prenese v skupino -sh cisteina V β-cetoacil-ACP sintazi kompleksa reakcija, ki jo katalizira encim acetil-CoA-ACP transacetila (AT) (AT).
Nato se malonska skupina prenese iz molekule malonil -CoA v skupino -SH skupine ACILO Group (ACP), ki je del kompleksne maščobne kisline.
Zaporedje štirih reakcij za vsak "vrnitev" reakcijskega cikla je naslednje:
- Kondenzacija: Acetilne in malonilne skupine, "naložene" v encim. V tem koraku je molekula CO2 in jo katalizira sintaza β-zoacil-ACP (acetilna skupina zavzema položaj „končnega metila“ kompleksa acetoacetil-ACP).
- Zmanjšanje karbonilne skupine: karbonilna skupina v položaju C3 acetoacetil-ACP se zmanjša na tvorbo D-β-hidroksibutiril-ACP, reakcija, ki jo katalizira β-cetoacil-ACP reduktaza, ki jo NADPH uporablja kot elektronski donor.
- Dehidracija: C2 in C3 ogljikovi ogljikom D-β-hidroksibutiril-ACP so brez molekul vode, ki tvorijo dvojno vez, ki se konča s proizvodnjo nove spojine trans-∆2-butenoil-ACP. Ta postopek posreduje encim β-hidroksiacil-ACP dehidrataze (HD) (HD).
- Zmanjšanje dvojne vezi: dvojna vez spojine, ki nastane v koraku dehidracije, je nasičena (zmanjšana), da se povzroči butiril-ACP z reakcijo, ki jo katalizira encim jeza-ACP reduktaza (ER), ki uporablja tudi NADPH kot reducirajoče sredstvo.
Reakcije sinteze se pojavijo, dokler ne nastane molekula palmitata (16 ogljikovih atomov), ki se hidrolizira iz encimskega kompleksa in se sprošča kot možen predhodnik maščobnih kislin večjih dolžinskih verig, ki jih proizvajajo raztezanje sistemov maščobnih kislin, ki se nahajajo v gladkem del endoplazemskega retikuluma in v mitohondrijih.
Lahko vam služi: favna in flora perujskega morjaDruge spremembe, ki jih te molekule lahko trpijo.
- Sinteza eikosanoidov
Eikosanoidi so celični lipidi, ki imajo funkcije, kot so molekule messengerja s kratko območje, ki jih proizvajajo nekatere tkanine za komunikacijo s sosednjimi tkaninami. Te molekule so sintetizirane iz polinenasičenih maščobnih kislin 20 ogljikovih atomov.
Prostaglandini
Kot odgovor na hormonski dražljaj encim fosfolipaza napade membranske fosfolipide in sprošča arakidonato iz glicerola ogljika 2. Ta spojina se pretvori v prostaglandine, zahvaljujoč encimu gladkega endoplazemskega retikuluma z bifunkcionalno aktivnostjo: ciklooksigenaza (COX) ali prostaglandin H2 sintaza.
Tromboxan
Prostaglandine lahko pretvorimo v tromboksi, zahvaljujoč tromboksanom, sintažnim v krvnih trombocitih (trombociti). Te molekule sodelujejo v začetnih korakih koagulacije krvi.
- Sinteza triacilglicerolov
Maščobne kisline so temeljne molekule za sintezo drugih bolj zapletenih spojin v celicah, kot so triacilgliceroli ali lipidni glicerofosfolipidni lipidni membrani (procesi, ki so odvisni od celičnih presnovnih potreb).
Živali proizvajajo triacilglicerole in gliceofosfalipide iz dveh pogostih predhodnikov: maščobni acil in 3-fosfat L-glicerol. Acil maščob-CoA nastajajo s sintezami acil-CoA, ki sodelujejo pri β-oksidaciji, medtem ko 3-fosfat L-glicerol dobimo iz glikolize in z delovanjem dveh alternativnih encimov: 3-fosfat glicerol dehidrogenaza in kinazni glicerolol.
Triacilglicerole tvorijo reakcijo med dvema molekulama Flo-CoA acil in 3-fosfatnim diacilglicerol molekuli; Te prenosne reakcije katalizirajo s posebnimi transfernami.
V tej reakciji je sprva fosfatidna kislina, ki jo defosforilira z encimsko fosfatidno kislinsko fosfatazo, da proizvede 1,2-diacilglicerol, ki je spet sposoben sprejeti tretjo molekulo maščobnega acila, ki proizvaja triacilglyceroll.
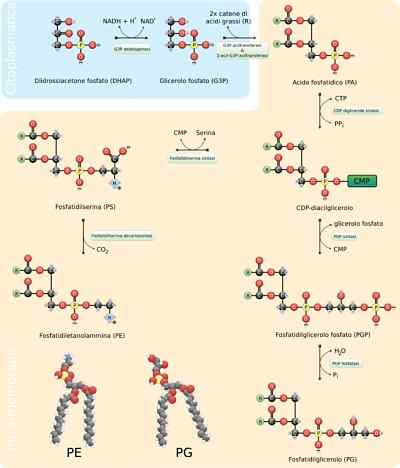
- Sinteza fosfolipidov
Fosfolipidi so izjemno spremenljive molekule, saj se veliko različnih lahko tvorijo s kombinacijo maščobnih kislin in "glave" skupin z glicerolskim okostjem (gliceofosfolipidi) ali sfinozinom (sfinolipidi).
Generalna sklop teh molekul zahteva sintezo okostja glicerola ali sferičnega združenja z ustreznimi maščobnimi kislinami, bodisi s sterifikacijo ali amidacijo, dodajanje hidrofilne skupine "glave" skozi fosfodirsko vez in, če je potrebno , sprememba ali izmenjava teh zadnjih skupin.
V evkariotih se ta proces pojavlja v gladkem endoplazemskem retikulumu in tudi v notranji mitohondrijski membrani, kjer lahko ostanejo v nedogled ali od koder jih je mogoče premestitve v druge kraje.
Reakcijski koraki
Prvi koraki sinteze reakcije glicerofosfolipidov so enakovredni tistim iz proizvodnje triacilglicerolov, saj je molekula 3-fosfata glicerola esterificirana na dve molekuli maščobnih kislin v karbonih 1 in 2, kar tvori fosfatidno kislino. Običajno je najti fosfolipide, ki imajo nasičene maščobne kisline v C1 in nenasičene v C2 glicerola.
Vam lahko služi: uporFosfatidno kislino lahko proizvajamo tudi s fosforilacijo molekule diacilglicerola, ki je že sintetizirana ali "reciklirana".
Polarne skupine "glave" teh molekul se tvorijo s fosfodiésterskimi povezavami. Prva stvar, ki bi se morala zgoditi, da je ta postopek pravilno, je "aktivacija" ene od hidroksilnih skupin, ki sodeluje v postopku s pomočjo hidroksila, ki sodeluje v reakciji.
Če se ta molekula veže na diacilglicerol, se tvori CDP-diacilglicerol ("aktivirana" oblika fosfatidne kisline), vendar se to lahko zgodi tudi v hidroksilni skupini skupine "glava".
V primeru fosfatidilserina se na primer diacilglicerol aktivira s kondenzacijo molekule fosfatidne kisline s trif-citidin citidin molekulo (CTP), ki tvori CDP-diacilglicerol in izloči pirofosfat.
Če se molekula MMP (citidin monofosfat) premika skozi nukleofilni napad serina ali hidroksilnega hidroksila v ogljiku 1 3-fosfatnega glicerola. Fosfat proizvaja fosfatidilglicerol.
Obe molekuli, proizvedeni tako.
- Sinteza holesterola
Holesterol je bistvena molekula za živali, ki jih lahko sintetizirajo njegove celice, zato v vsakodnevni prehrani ni bistvenega pomena. Ta molekula 27 ogljikovih atomov nastaja iz predhodnika: acetat.
Ta kompleksna molekula je nastala iz acetil-CoA v štirih glavnih fazah:
- Kondenzacija treh acetatnih enot za tvorbo mevalonato, vmesna molekula 6 ogljikov (najprej acetoacetil-CoA molekula nastane z dvema acetil-CoA (tiolaznim encimom) in nato še eno β-hidroksi-β-metilglaril-CoA CoA) (encim HMG-CoA Syntheta). Mevalonato je oblikovan iz HMG-CoA in zahvaljujoč encimu HMG-CoA reduktaze.
- Pretvorba mevalonata v izoprenske enote. Prve 3 fosfatne skupine se prenesejo iz 3 molekul ATP v Mevalonato. Eden od fosfatov je izgubljen skupaj s sosednjo karbonilno skupino in je oblikovan
- Polimerizacija ali kondenzacija 6 izoprenskih enot 5 ogljikovih atomov, da tvorimo 30 -ogljikove atome (linearna molekula) britje (linearna molekula).
- Cicracija escuala, da tvori 4 obroče steroidnega jedra holesterola in kasnejše kemične spremembe: oksidacije, migracija in izločanje metilnih skupin itd., Kaj daje holesterol.
Reference
- Garrett, r. H., & Grisham, c. M. (2001). Načela biokemije: s človeškim fokusom. Založba Brooks/Cole.
- Murray, r. K., Granner, d. K., Mayes, str. Do., & Rodwell, v. W. (2014). Harperjeva ilustrirana biokemija. McGraw-Hill.
- Nelson, d. L., Lehninger, a. L., & Cox, m. M. (2008). Lehningerjeva načela biokemije. Macmillan.
- Jacquemyn, j., Cascaho, a., & Goodchild, r. In. (2017). INS in izhodi endoplazemske retikulum - nadzorovana biosinteza lipidov. Poročila EMBO, 18 (11), 1905-1921.
- Ohlrogge, j., & Brskaj, J. (devetnajst devetdeset pet). Lipidna biosinteza. Rastlinska celica, 7 (7), 957.
- « José López Portillo Biografija, vlada in prispevki
- Tropska džungla v mehiški značilnosti, lokacija, relief, flora, favna »