

Mehanizmi za razmnoževanje DNK v prokariotih in evkariotih
- 4177
- 812
- Ignacio Barrows
The Podvajanje DNK (Desoxyribonucleic kislina) Sestavljen je iz kopiranja genoma, torej vseh genetskih informacij, ki jih vsebuje DNK organizma, da bi ustvarili dve enaki kopiji. Genom ima potrebne informacije za izgradnjo celotnega organizma.
Pred delitvijo celic se pojavi podvajanje DNK. Skozi mejozo se pojavijo gamete za spolno razmnoževanje. Z mitozo pride do zamenjave celic (na primer koža in kri) in razvoj (na primer tkiva in organi).
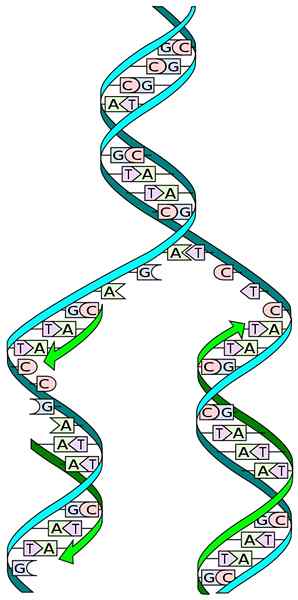
 Vir: jaz, Madprime [CC0]
Vir: jaz, Madprime [CC0] Poznavanje strukture DNK nam omogoča, da razumemo način, kako se pojavi njena replikacija. Struktura DNK je sestavljena iz dvojnega propelerja, sestavljenega iz dveh antipaallalnih verig zaporednih nukleotidov, katerih dušikove baze so posebej dopolnjene.
Med replikacijo vsaka veriga dvojne verige DNK deluje kot kalup za biosintezo nove verige. Dve na novo sintetizirani verigi imata podlage, ki se dopolnjujeta z osnovami verige plesni: adenin (a) s timino (t) in citozin (c) z gvaninom (g).
Pri podvajanju DNK sodeluje več encimov in beljakovin. Na primer, odpiranje dvojnega propelerja DNK, ohranjanje DNK odprt in dodajanje deoksiribonukleosidos-5'-trifosfata (DNTP) za tvorbo nove verige.
[TOC]
Podvajanje DNK je polkonservativno
Na podlagi strukture DNK sta Watson in Crick predlagala, da se razmnoževanje DNK pojavi na polkonzervativen način. To sta pokazala Meselson in Stahl z oznako DNK Escherichia coli S težkim izotopom dušika, petnajstN, za več generacij vzorec porazdelitve v kulturnem mediju s svetlobnim dušikom, 14N.
Messelson in Stahl sta ugotovila, da sta v prvi generaciji molekula DNK hčere imela, vsako molekulo, označeno z verigo s težkim dušikovim izotopom in drugo s svetlobnim izotopom. Za razliko od starševske molekule DNK, ki sta imeli dve verigi označeni s težkim izotopom, petnajstN.
V drugi generaciji je bilo 50% molekul DNK podobno kot prve generacije, ostalih 50%. Razlaga tega rezultata je, da ima dvojna hči propeler starševsko verigo (ki deluje kot kalup) in novo verigo.
Mehanizem polkonzervativnega razmnoževanja pomeni ločitev verig DNK in parjenje komplementarnih baz z zaporednimi nukleotidi, ki proizvajata dve hčerki hčere z dvojnimi hčerkama.
Podvajanje v baterijah
Začetek razmnoževanja DNK v bakterijah
DNK bakterij je sestavljen iz krožnega kromosoma in ima samo eno mesto izvora podvajanja. Na tem mestu se biosinteza obeh hčerk zgodi dvosmerno in tvori dve replikacijski vilici, ki se premikata v navodilih, ki nasprotujejo izvoru. Na koncu so vilice, dokončajo podvajanje.
Replikacija se začne z zvezo beljakovin DNAA na mestu izvora. Ti beljakovine tvorijo kompleks. Potem so proteini Hu in IHF vezani.
Nato se pridružijo proteini DNAC, zaradi česar se helikoze DNK pridružijo. Ti pomagajo sprostiti DNK in razbiti vodikove vezi, nastale v baznih parih. Torej, dve verigi se še bolj ločita in tvorita dve preprosti verigi.
Vam lahko služi: kaj je kromosomska permutacija?Topoisomeraza II ali DNA girasa se premika pred DNK hemicic. Proteini vezave DNK z eno verigo (SSB) ohranjajo ločene verige DNA. Tako se lahko začne biosinteza hčerinske verige.
Biosinteza verig DNK hčere v bakterijah
Prima Encim je odgovoren za sintezo kratkih verig RNA, imenovanih prajmeri, ki imajo dolžine od 10 do 15 nukleotidov. DNK polimeraza začne dodajati 5'-trifosfat (DNTPS) 3'-OH sladkornega sladkorja, po katerem veriga še naprej raste na istem koncu.
Ker so verige DNK protiparalne, se v vodilni verigi sintetizira temeljni premaz in veliko temeljnih premazov v zapozneli verigi. Zaradi tega je biosinteza zapoznele verige prekinjena. Čeprav so verige DNK protiparalne, se vilice za razmnoževanje premikajo v eno smer.
DNK polimeraza je odgovorna za tvorbo kovalentnih vezi med sosednjimi nukleotidi novo sintetiziranih verig v smeri 5'®3 '. V In. coli, Obstaja pet DNK polimeraze: DNK polimeraze I in III izvajajo podvajanje DNK; in DNK polimeraze II, IV in V so odgovorni za ponovitev in ponovitev poškodovane DNK.
Večino podvajanja naredi DNK polimeraza III, ki je holoencim, ki ima 10 različnih podenot z več funkcijami pri podvajanju DNK. Na primer, podenota Alpha je odgovorna za izdelavo vezi med nukleotidi.
Encimski kompleks je odgovoren za razmnoževanje DNK v bakterijah
Helikaza DNK in prima se združujeta v kompleks, imenovan Primosoma. To se premika vzdolž DNK in deluje usklajeno, da ločita obe starševski verigi, sintetizirajo temeljne premaze vsak določen interval na zakasnjeni verigi.
Primosome se fizično veže na DNA polimerazo III in tvori ponovljiv. Dve DNK polimerazi III sta odgovorni za razmnoževanje DNK vodnika in zapoznele verige. Glede na DNA polimerazo III zakasnjena veriga tvori zanko, kar omogoča dodajanje nukleotidov v to verigo v isti smeri vodilne verige.
Dodajanje nukleotidov v vodilno verigo je neprekinjeno. Medtem ko je v zamudi prekinjen. Fragmenti 150 nukleotidov dolžine so oblikovani, imenovani Okazaki fragmenti.
Aktivnost eksonukleaze 5 ' -> 3' DNA polimeraze I je odgovorna za odstranjevanje prajmov in polnjenje, dodajanje nukleotidov. Encimska ligaza tesni vrzeli med fragmenti. Podvajanje se konča, ko sta dva replikacijska hoquilla v zaporedju dokončanja.
Protein se vaš veže na zaporedje zaključka in ustavi gibanje vilic za podvajanje. Topoisomeraza II omogoča ločitev obeh kromosomov.
Desoxiribonukleotidi Triphosfate uporablja DNA polimeraza
Deksinukleozid trifosfat (DNTP) vsebuje tri fosfatne skupine, združene z ogljikom 5 'deoksiribosa. DNTPS (DATP, DTTP, DGTP in DCTP) se pridružijo verigi plesni po pravilu AT/GC.
Vam lahko služi: plesiomorphyDNA polimeraza katalizira naslednjo reakcijo: hidroksilna skupina (-OH) 3 'nukleotida rastoče verige reagira z alfa fosfatom dohodnega DNTP, sprošča anorganski pirofosfat (PPI) (PPI). Hidroliza PPI proizvaja energijo za tvorbo kovalentne vezi ali fosfodiésterske vezi med nukleotidi rastoče verige.
Mehanizmi, ki zagotavljajo zvestobo podvajanja DNK
Med podvajanjem DNK polimeraza III DNK naredi napako na 100 milijonov nukleotidov. Čeprav je verjetnost napake zelo nizka, obstajajo mehanizmi, ki zagotavljajo zvestobo pri podvajanju DNK. Ti mehanizmi so:
1) stabilnost v osnovnem aparju. Energija vodikove vezi med AT/GC je večja kot v napačnih baznih parih.
2) Struktura aktivnega mesta DNA polimeraze. DNK polimeraza prednostno katalizira nukleotide s pravilnimi osnovami v nasprotni verigi. Slabo parjenje baz povzroči izkrivljanje dvojnega propelerja DNA, kar preprečuje, da bi napačni nukleotid zasedel aktivno mesto encima.
3) Test branja. DNK polimeraza identificira vključene napačne nukleotide in jih odpravi iz hčerinske verige. Aktivnost eksonukleaze polimerazne DNK razbije fosfodiésterjeve vezi med nukleotidi na 3 'koncu nove verige.
Podvajanje DNK v evkariotih
Za razliko od razmnoževanja v prokariotih, katerih razmnoževanje se začne na enem mestu, se evkariontska replikacija začne na več mestih izvora in vilice za razmnoževanje se dvosmerno premika. Kasneje so združene vse replikacijske vilice, ki tvorijo dve sestrski kromatidi, ki sta se pridružili v centromeru.
Eukaryotas ima veliko vrst DNK polimeraze, katerih imena uporabljajo grške črke. DNK polimeraza α tvorijo kompleks s primo. Ta kompleksna sintetizira kratke prajmese, sestavljene iz 10 nukleotidov RNA, ki jim sledi 20 do 30 DNK nukleotidov.
Nato DNK polimeraza ε tudi δ katalizira raztezanje hčerinske verige iz temeljnega premaza. DNK polimeraza ε Vključen je v sintezo vodilne verige, medtem ko polimeraza DNA δ Sintetizirajte zapoznelo verigo.
DNK polimeraza δ Nadaljujte levi fragment Okazakija, dokler ne dosežete desnega temeljnega premaza RNA in ustvarite kratek dvig temeljnega premaza ("kratka loputa"). Za razliko od prokariotov, kjer polimerazna DNK izloči temeljni premaz, v evkariotih encim encima endonukleaze odpravlja temeljni premaz RNA.
Nato DNK ligaza tesnilo fragmente DNK, ki so sosednji. Dokončanje podvajanja poteka z disociacijo beljakovin za razmnoževanje.
The Podvajanje DNK v evkariotih in celičnem ciklu
Podvajanje v evkariotih se pojavi v fazi S celičnega cikla. Ponovne molekule DNK so med mitozo ločene v dveh hčerinskih celicah. Faza G1 in G2 ločita fazo in mitozo S. Napredek skozi vsako fazo celičnega cikla močno regulira kinaze, fosfataze in proteaze.
V fazi G1 celičnega cikla se kompleks za prepoznavanje izvora (OCR) pridruži mestu izvora. To spodbudi zvezo MCM helicas in drugih beljakovin, kot sta CDC6 in CDT1. LAS Helicase MCM se pridruži verigi Guide.
Vam lahko služi: povezani geniV fazi S postane PerC aktivno mesto za razmnoževanje. Proteini OCR, CDC6 in CDT1 se sprostijo, MCM helikaza pa se premika v smeri 3 'do 5'. Ko se replikacija konča, se bo to znova zagnalo v naslednjem celičnem ciklu.
Podvajanje koncev kromosomov v evkariotih
Konci kromosomov so znani kot telomeri, ki so sestavljeni iz ponavljajočih se zaporedja v tandemu in 3 'regije, ki se odlikuje, od 12 do 16 nukleotidov dolžine.
DNK polimeraza ne more ponoviti 3 'konca verig DNK. To je zato, ker lahko DNK polimeraza sintetizira DNK v naslovu 5'-3 'in lahko samo podaljša že obstoječe verige, ne da bi v tej regiji sintetiziral temeljni premaz. Posledično se telomeri skrajšajo v vsakem krogu podvajanja.
Encim telomeraze preprečuje skrajšanje telomerov. Telomeraza je encim, ki ima beljakovinske in RNA podenote (tretji). Slednje se veže na večkratne sekvence DNK in omogoča telomerazo, da se pridruži 3 'konec telomera.
Zaporedje RNA za Unijo, ki se nahaja kot kalup za sintezo zaporedja šestih nukleotidov (polimerizacija) na koncu verige DNK. Raztezanje telomera katalizira podenote telomeraze, imenovano reverzno telomerazno transkriptazo (TERT).
Po polimerizaciji poteka translokalizacija, ki je sestavljena iz gibanja telomeraze na nov konec verige DNK, ki se do konca pridruži šest drugih nukleotidov.
Funkcije drugih DNK polimeraz v evkariotih
DNK polimeraza β Ima pomembno vlogo pri izločanju napačnih baz DNK, vendar ni vključen v podvajanje DNK.
Številni odkrili DNK polimeraze spadajo v skupino polimeraze, ki rešuje translezijo. Te polimeraze so odgovorne za sintezo dopolnilnih verig v poškodovanem območju DNK.
Obstaja več vrst polimeraz, ki "prenašajo translezijo". Na primer DNK polimeraza η Lahko ponovite o Timini dímeros, ki ga proizvaja UV svetloba.
Podvajanje DNK v arheobakteriji
Podvajanje DNK arheobakterije je podobno tistemu, ki je naveden v evkariotih. To je posledica naslednjega: 1) beljakovine, ki sodelujejo pri podvajanju, so bolj podobne evkarioti kot pri prokarioti; in 2) Čeprav obstaja samo eno mesto za razmnoževanje kot v prokarioti, je njegovo zaporedje podobno mestu evkariontskega izvora.
Podobnost pri razmnoževanju med loki in evkarioti podpira idejo, da sta obe skupini filogenetsko bolj povezani med seboj kot kateri koli od njih s prokarioti.
Reference
- Brooker, r. J. 2018. Analiza genetike in načela. McGraw-Hill, New York.
- Hartwell, l. H., Goldberg, m. L., Fischer, J. Do., Kapuca, l. 2018. Genetika - iz genov genov. McGraw-Hill, New York.
- Kušić-Tišma, j. 2011. Temeljni vidiki podvajanja DNK. INTECH Open Access, Hrvaška.
- Lewis, r., 2015. Koncepti in aplikacije človeške genetike. McGraw-Hill, New York.
- Pierce, b. Do. 2005. Genetika - konceptualni pristop. W. H. Freeman, New York.