

Značilnosti, funkcije, funkcije, komponente
- 2130
- 316
- Lee Farrell
On jedrska pora (Iz grščine, pore = korak ali tranzit) je "vrata" jedra, ki omogoča večino prevoza med nukleoplazmo in citoplazmo. Jedrska pore se pridruži notranjim in zunanjim membranam jedra, da tvori kanale, ki služijo za transport beljakovin in RNA.
Beseda Poro ne odraža zapletenosti zadevne strukture. Zaradi tega se je bolje sklicevati na kompleks jedrskih pore (CPN), namesto na jedrsko pore. CPN lahko doživi spremembe v svoji strukturi med prevozom ali stanja celičnega cikla.
V zadnjem času je bilo ugotovljeno, da imajo nukleoporini, beljakovine, ki sestavljajo CPN, pomembno vlogo pri uravnavanju izražanja genov. Ko se pojavijo mutacije, ki vplivajo na delovanje nukleoporinov, se patologije proizvajajo pri ljudeh, kot so avtoimunske, kardiomiopatije, virusne okužbe in rak.
[TOC]
Značilnosti
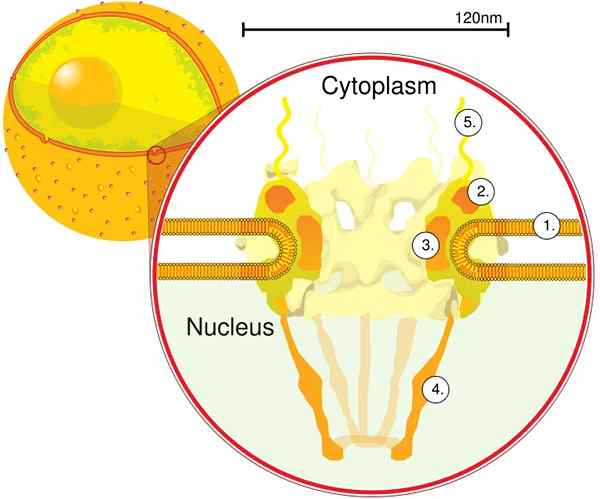
Skozi elektronsko tomografijo je bilo ugotovljeno, da ima CPN debelino 50 nm, zunanji premer med 80 in 120 nm in notranji premer 40 nm. Velike molekule, kot je velika podenota ribosomov (PM 1590 kDa), lahko izvozimo zunaj jedra skozi CPN. Ocenjuje se, da je med 2000 in 4000 CPN na jedro.
Molekulska masa posameznega CPN je približno med 120 in 125 MDA (1 MDA = 106 Da) v vretenčarjih. V nasprotju s tem je CPN manjši v kvasovkah, v katerih ima približno 60 MDA. Kljub ogromni velikosti CPN so nukleoporini v vseh evkariotih zelo ohranjeni.
Translokalizacija skozi CPN je hiter proces, katerega hitrost je 1000 translokacij/sekundo. Vendar CPN ne določa smeri samega transportnega toka.
To je odvisno od gradienta Rangtp, ki je v jedru večje kot v citosolu. Ta gradient vzdržuje faktor gvaninskega izmenjevalnika.
Med celičnim ciklom CPN -ji doživljajo cikle montaže in razsodnosti. Sklop se zgodi na vmesniku in takoj po mitozi.
Funkcije
Ribonukleinska kislina (majhna jedrska RNA, messenger RNA, prenos RNA), beljakovine in ribonukleoproteini (RNP) je treba aktivno prevažati prek CPN. To pomeni, da je potrebna energija hidrolize ATP in GTP. Vsaka molekula se prevaža na določen način.
Na splošno so molekule RNA napolnjene z RNP kompleksi, ki tvorijo beljakovine, ki se na ta način izvažajo. V nasprotju s tem morajo biti beljakovine, ki se aktivno prevažajo v jedro.
Vam lahko služi: mezosomBeljakovine, ki se izvažajo v jedro, morajo imeti izvozni signal v jedro (NES), bogato z levcinsko aminokislino.
Poleg tega, da olajšajo transport med jedrom in citoplazmo, so CPN vključeni v organizacijo kromatina, regulacijo genske ekspresije in popravilo DNK. Nukleoporini (NUP) spodbujajo aktivacijo ali zatiranje transkripcije, ne glede na stanje širjenja celic.
V kvasovkah se nudi v CNP jedrskega zavijanja. V metazoarjih so v notranjosti. Opravljajo iste funkcije v vseh evkariotih.
Uvoz snovi
Skozi CPN je pasivno razširjanje majhnih molekul v tako smeri kot v aktivnem transportu, uvozu beljakovin, izvozu RNA in ribonukleoproteinov (RNP) ter dvosmernih molekul. Slednje vključuje RNA, RNP in beljakovine, ki sodelujejo pri signalizaciji, biogenezi in zamenjavi.
Uvoz beljakovin znotraj jedra se zgodi v dveh korakih: 1) vezava beljakovin na citoplazemsko stran CPN; 2) ATP odvisna translokalizacija prek CPN. Ta postopek potrebuje hidrolizo ATP in izmenjavo GTP/BDP med jedrom in citoplazmo.
Po prometnem modelu se kompleks sprejemanja in beljakovin premika po kanalu skozi Union, disociacijo in zvezo znova na večkratne FG zaporedja nukleoporinov. Na ta način se kompleks premika iz enega nukleoporina v drugega znotraj CPN.
Izvoz snovi
Je podoben uvozu. Ran GTPase nalaga usmeritev za prevoz skozi CNP. RAN je molekularno stikalo z dvema konformacijskima stanjem, odvisno od tega, ali je povezan z BDP ali GTP.
Dva specifična regulativna proteina sprožita pretvorbo med obema stanje: 1) citosolni protein, ki aktivira citosolno GTPazo (GAP), ki proizvaja hidrolizo GTP in tako pretvori Ran-GTP v Ran-GDP; in 2) jedrski Guaninski menjalni faktor (GEF), ki spodbuja izmenjavo BDP z GTP in pretvori Ran-GDP v RAN-GTP.
Citosol v glavnem vsebuje Ran-GDP. Jedro vsebuje predvsem RAN-GTP. Ta gradient obeh konformacijskih oblik Ran usmerja prevoz v ustreznem naslovu.
Uvoz sprejemnika skupaj s položajem olajša ponovitev FG. Če pridete do jedrske strani CNP, se RAN-GTP pridruži sprejemniku, da sprosti vaš položaj. Tako RAN-GTP ustvarja smer uvoza.
Jedrski izvoz je podoben. Vendar RAN-GTP v jedru spodbuja zvezo položaja do izvoznega sprejemnika. Ko se izvozni sprejemnik premakne skozi pore proti citosolu, se sreča z Ran-GAP, kar sproži hidrolizo GTP na BDP. Končno se sprejemnik sprosti s svojega položaja in v citosolu RAN-GDP.
Vam lahko služi: polimorfonuklearne levkocitePrevoz RNA
Izvoz nekaterih vrst RNA je podoben izvozu beljakovin. Na primer, ARNT in RNAN (majhno jedrsko) uporabljata gradient RangtP in se s pomočjo prenašata skozi CPN. Izvoz zrelih ribosomov je odvisen tudi od gradienta RangtP.
MRNA se izvaža na zelo drugačen način kot beljakovine in druge RNA. Za svoj izvoz mRNA tvori kompleks RNP Messenger (RNPM), v katerem je molekula RNM obdana s stotimi beljakovinskimi molekuli. Ti beljakovine imajo obdelavo, Zakrivanje, Empalme in poliadentilly MNA.
Celica mora biti sposobna razlikovati med RNPM z zrelim RNM in RNPM z nezrelo mRNA. MRNA, ki tvori kompleks RPNM, bi lahko sprejela topologije, ki jih je treba preoblikovati za prevoz. Preden RNPM vstopi v CPN, se pojavi kontrolni korak, ki ga izvedejo TRAMP in eksozomski proteinski kompleksi.
Ko je zrel RNPM sestavljen, se RPNM prevaža skozi kanal s transportnim sprejemnikom (NXF1-NXT1). Ta sprejemnik potrebuje hidrolizo ATP (ne na gradient RangtP), da ugotovi preureditev usmerjenosti RNPM, ki bo dosegla citoplazmo.
Kompleks jedrskih por in nadzor izražanja genov
Nekatere študije kažejo, da lahko komponente CPN vplivajo na regulacijo genetske ekspresije s pomočjo kontrole kromatinske strukture in njegovo dostopnost do transkripcijskih faktorjev.
V evkariotih novejše evolucije je heterokromatin prednostno nameščen na obodu jedra. To ozemlje prekinejo evkromatinski kanali, ki jih vzdržuje jedrska košara CPN. Povezava jedrske košare z evkromatinom je povezana s transkripcijo genov.
Na primer, aktiviranje transkripcije na ravni CPN pomeni interakcijo komponent jedrske košare z beljakovinami, kot sta histon acetiltransferaza saga in izvozni faktorji RNA.
Tako je jedrska košara platforma za številne vzdrževalne gene (Gospodinjstvo) visoko prepisani in geni, ki jih močno povzročijo spremembe v okoljskih razmerah.
Jedrski pore in virološki kompleks
Okužba z virusno evkariotsko celico je odvisna od CPN. V vsakem primeru virusne okužbe je njegov uspeh odvisen od DNK, RNA ali RPN, ki gredo skozi CPN, da dosežejo svoj končni cilj, to je razmnoževanje virusa.
Vam lahko služi: citoplazemski vključki: kaj so, značilnosti, funkcijeVirus APE 40 (SV40) je bil eden najbolj preučenih modelov, ki je preučil funkcijo CPN v translokalizaciji znotraj jedra. To je zato, ker ima SV40 majhen genom (5.000 baz).
Pokazalo se je, da je transport virusne DNK olajšan beljakovinski proteini virusa, ki ščitijo virus, dokler jedro ne doseže.
Komponente
CPN je vgrajen v jedrsko ovojnico in je sestavljen iz približno 500 do 1000. Ti proteini so organizirani v strukturnih podkompleksih ali modulih, ki medsebojno delujejo.
Prvi modul je osrednja komponenta ali obroč znotraj pore v obliki peščene ure, ki je omejena s še enim obročem s premerom 120 nm na obeh straneh, intranuklearni in citoplazemski. Drugi modul so jedra in citoplazma obroči (premera 120 nm), ki se nahajajo okoli komponente v obliki črke peska.
Tretji modul je osem filamentov, ki se projicirajo iz 120 nm obroča znotraj nukleoplazme in tvori strukturo v obliki košare. Četrti modul je sestavljen iz filamentov, ki se projicirajo na stran citoplazme.
Kompleks, ki ga oblikuje Y, sestavljen iz šestih NUP -jev in SEH 1 in Sec 13 proteinov, je največji in najbolje značilen kompleks CNP. Ta kompleks je bistvena enota, ki je del odrov CPN.
Kljub nizki podobnosti med zaporedji NUPS je oder CPN zelo ohranjeno v vseh evkariotih.
Reference
- Beck, m., Poškodovan, e. 2016. Kompleks jedrskih por: razumevanje njegove funkcije s strukturnim vpogledom. Ocene narave, biologija molekulskih celic, doi: 10.1038/nrm.2016.147.
- Ibarra, a., Hetzer, m.W. 2015. Jedrski beljakovine pore in nadzor funkcij genoma. Geni in razvoj, 29, 337-349.
- Kabachinski, g., Schwartz, t.Ali. 2015. Kompleks jedrskih por - struktura in delovanje na prvi pogled. Journal of Cell Science, 128, 423–429.
- Knocknhauer, k.In., Schwartz, t.Ali. 2016. Kompleks jedrskih por kot prilagodljiva in dinamična vrata. Cell, 164, 1162-1171.
- PTAK, c., Aitchison, J.D., Wozniak, r.W. 2014. Večfunkcionalno jedrsko s kompleksom: platforma za nadzor izražanja genov. Trenutno mnenje o celični biologiji, doi: 10.1016/j.CEB.2014.02.001.
- Stawicki, s.Str., Steffen, j. 2017. Republikacija: kompleks jedrskih por - celovit pregled strukture in funkcije. International Journal of Academic Medicine, 3, 51–59.
- Tran, e.J., Wente, s.R. 2006. Dinamično jedrsko s kompleksi: Življenje na robu. Celica, 125, 1041-1053.