

Značilnosti mikrofilamentov, struktura, funkcije, patologija
- 1325
- 225
- Roman Schamberger
The mikrofilamente o aktinske filamente, so ena od treh temeljnih komponent citoskeleta evkariontskih celic.
V evkariotih so geni, ki kodirajo aktinske mikrofilamente, v vseh organizmih zelo ohranjeni, zato se pogosto uporabljajo kot molekularni markerji za različne študije.
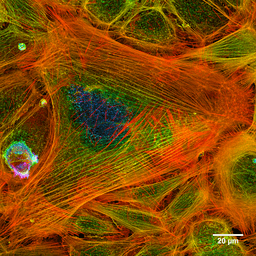
Fotografija aktinskih filamentov barvane celice (vir: Howard Vindin [CC BY-SA 4.0 (https: // creativeCommons.Org/licence/by-sa/4.0)] prek Wikimedia Commons) Mikrofilamente se porazdelijo po celotnem citosolu, vendar so v osnovnem območju še posebej v plazemski membrani, kjer tvorijo zapleteno mrežo in so povezani z drugimi posebnimi beljakovinami, da tvorijo citoskelet.
Mikrofilamentne mreže v citoplazmi celic sesalcev kodirata dva od šestih genov, opisanih za aktin, ki so vključeni v dinamiko mikrofilamentov in so med diferenciacijo matičnih celic celo zelo pomembne.
Številni avtorji se strinjajo, da so mikrofilamenti najbolj raznoliki, vsestranski in pomembni proteini citoskeleta večine evkariontskih celic, zato je treba spomniti, da jih ne najdemo v prokariotskih mikroorganizmih.
V tej vrsti celic pa so mikrofilamente homologne filamente, ki jih tvori drug protein: MREB protein.
Trenutno se misli, da je gen, ki kodira ta protein, možen gen prednikov za evkariontski aktin. Vendar je homologija aminokislinskih zaporedja, ki tvori protein MREB, le 15% glede na zaporedje aktina.
Kot je temeljni del citoskeleta, lahko vsaka fenotipska okvara tako v mikrotubulah kot v vmesnih nitkah in aktinskih mikrofilamentih (citoskelet) povzroči različne celične in sistemske patologije.
[TOC]
Značilnosti in struktura
Mikrofilamente sestavljajo beljakovinske monomere družine aktina, ki so v evkariontskih celicah izjemno obilni kontraktilni proteini, saj sodelujejo tudi pri krčenju mišic.
Te nitke imajo premer med 5 in 7 nm, zato so znane tudi kot tanke nitke in so sestavljene iz dveh oblik aktina: kroglične oblike (aktin g) in nitaste oblike (aktin f).
Beljakovine, ki sodelujejo v citoskeletu.
Vam lahko služi: citoplazma: funkcije, deli in značilnostiDelež globularnega aktina in filamentnega aktina v citosolu je odvisen od celičnih potreb, saj so mikrofilamenti zelo spremenljive in vsestranske strukture, ki sta nenehno rasti in skrajšajo zaradi polimerizacije in depolimerizacije.
Actin G je majhen kroglični protein, sestavljen iz skoraj 400 aminokislin in približno 43 kDa molekulske mase.
Aktinski monomeri, ki sestavljajo mikrofilamente.
Actin g je povezan z molekulo Ca2+ in drugim ATP, ki stabilizira njegovo kroglično obliko; Medtem ko aktin F dobimo po hidrolizi terminalnega fosfata molekule ATP v aktinu G, ki prispeva k polimerizaciji.
Organizacija
Aktinske filamente lahko organiziramo v obliki "žarkov" ali "omrežij", ki imajo v celicah različne funkcije. Snopi tvorijo vzporedne strukture, ki jih povezujejo precej togi križni mostovi.
Omrežja so na drugi strani bolj udobne strukture, kot so tri dimenzionalne mreže z lastnostmi pol -solidnih gelov.
Obstaja veliko beljakovin, ki so povezane z aktinskimi ali mikrofilamentnimi nitkami in so znane kot ABP (iz angleščine Actin vezavni proteini), ki imajo za to določena mesta.
Mnogi od teh beljakovin omogočajo, da mikrofilamente interakcijo z drugimi dvema komponentama citoskeleta: mikrotubule in vmesne filamente, pa tudi z drugimi komponentami notranjega obraza plazemske membrane.
Med drugimi beljakovinami, ki delujejo z mikrofilamini, so jedrski listi in spektrin (v rdečih krvnih celicah).
Kako se tvorijo aktinske filamente?
Ker kroglični aktinski monomeri se vedno pridružijo na enak način, usmerjeni v isto smer, imajo mikrofilamente določeno polarnost z dvema skrajnostma: ena "več" in ena "manj".
Polarnost teh filamentov je zelo pomembna, saj zaradi pozitivnega konca precej hitreje rastejo, kjer se dodajo novi aktin G monomeri.
 Grafični prikaz aktinskih mikrofilamentov (vir: Derivacijsko delo: Retama (pogovor) Thin_filament_Formation.SVG: Mikael Häggström [javna domena] prek Wikimedia Commons)
Grafični prikaz aktinskih mikrofilamentov (vir: Derivacijsko delo: Retama (pogovor) Thin_filament_Formation.SVG: Mikael Häggström [javna domena] prek Wikimedia Commons) Prva stvar, ki se zgodi med polimerizacijo aktinskih nitk.
Vam lahko služi: plazomolizaNa ta trimmer se obe konci dodata nove monomere, tako da filament raste. Aktinski monomeri lahko hidrolizirajo ATP z vsako zvezo, kar ima posledice hitrosti polimerizacije, saj deli Actin-ATP odpravljajo več težav kot Actin-ADP.
ATP ni potreben za polimerizacijo in betonska funkcija njegove hidrolize še ni razjasnjena.
Nekateri avtorji menijo, da so dogodki v polimerizaciji aktina hitri.
Uredba
Tako polimerizacija aktinskih filamentov kot tudi njena depolimerizacija sta procesi, ki jih visoko ureja niz specifičnih beljakovin, ki so odgovorni za preoblikovanje filamentov.
Primer beljakovin, ki uravnavajo depolimere. Drug protein, profilain, ima nasprotno funkcijo, saj spodbuja povezavo monomerov (s spodbujanjem ADP -jeve izmenjave za ATP).
Funkcije
Mikrofilamenti medsebojno delujejo z miozinskimi filamenti, ki so povezani s transmembranskimi beljakovinami, ki imajo domeno v citosolu in drugim v tujini, zato sodelujejo v procesih mobilnosti celic.
Ti mikrofilamenti, povezani s plazemsko membrano, posredujejo različne celične odzive na različne vrste dražljajev. Na primer, celično adhezijo epitelijskih tkiv usmerjajo transmembranski proteini, znani kot kadherina, ki sodelujejo z mikrofilamenti, da zaposlijo faktorje odziva.
Aktinske filamente medsebojno delujejo z vmesnimi filamenti, da se zunajcelični dražljaji prenašajo na ključna mesta, kot so ribosomi in kromosomi v jedru.
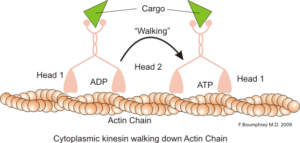 Predstavitev medcelične motorične funkcije aktinskih mikrofilamentov (vir: Boumphreyfr [CC by-SA 3.0 (https: // creativeCommons.Org/licence/by-sa/3.0)] prek Wikimedia Commons)
Predstavitev medcelične motorične funkcije aktinskih mikrofilamentov (vir: Boumphreyfr [CC by-SA 3.0 (https: // creativeCommons.Org/licence/by-sa/3.0)] prek Wikimedia Commons) Klasična in zelo preučena funkcija mikrofilamentov je njegova sposobnost oblikovanja "mostov", "tirnic" ali "avtocest" za gibanje miozinskega motoričnega proteina I, ki lahko nalagajo transportne vezikule iz organel na membransko plazmo na sekretorju poti.
Mikrofilamenti sodelujejo tudi z miozinom II, da vzpostavijo kontraktilni obroč, ki nastane med citokinezo, natančno med zadnjo stopnjo celične delitve, v kateri je citosol ločen od celic STEM in hčerine.
Vam lahko služi: polimorfonuklearne levkociteNa splošno mikrofilamenti v obliki aktina modulirajo porazdelitev nekaterih organel, kot so Golgijev kompleks, endoplazemski retikulum in mitohondriji. Poleg tega sodelujejo tudi v prostorskem pozicioniranju RNMS, tako da jih berejo ribosomi.
Celoten nabor mobilnih telefonov mikrofilamentov, zlasti tistih, ki so tesno povezani s plazemsko membrano, sodeluje pri tvorbi valovitih celičnih membran, ki imajo konstantno aktivno gibanje.
Sodelujejo tudi pri tvorbi mikrov in drugih skupnih izboklin na površini številnih celic.
Primer funkcij v jetrih
Mikrofilamenti sodelujejo v procesu izločanja žolča v hepatocitih (jetrnih celic) in tudi v peristaltičnih gibih (koordinirano krčenje) jetrnih kanalikulijev.
Prispevajo k diferenciaciji domen plazemske membrane, zahvaljujoč njihovi povezavi z različnimi citosolnimi elementi in nadzoru, ki ga izvajajo na topografiji teh znotrajceličnih elementov.
Povezane patologije
Malo je bolezni, povezanih s primarnimi napakami v strukturi ali z beljakovinami in regulativnimi encimi v sintezi mikrofilamentov, čeprav so te neposredno vključene v veliko število funkcij.
Nizek indeks bolezni in nepravilnosti v primarni strukturi mikrofilamentov je posledica dejstva, da na splošno obstaja več kodirajočih genov aktina in njegovih regulativnih beljakovin, pojav, znan kot "genetska odvečnost".
Ena najbolj preučenih patologij je vitrifikacija oocitov na njihovem citoskeletu, kjer opazimo prekinitev v kortikalni mikrofilamentni omrež.
Na splošno ta vitrifikacija povzroči kromosomsko disperzijo, saj vodi v nesoglasje pri zbiranju celotnega kromatina.
Celice, ki imajo večjo organizacijo in delež mikrofilamentov v svojem citoskeletu.
Okvarjeni ali atipični mikrofilamenti so bili povezani tudi z boleznijo kosti, znano kot pagetova bolezen.
Reference
- Aguilar-Cuenca, R., Llorente-González, c., Vicente, c., & Vicente-Manzanares, m. (2017). Dinamika adhezije, koordinirana z mikrofilamentom, poganja enocelične migracije in oblikuje tkiva. F1000Resarch, 6.
- Dva pravna sredstva, c. G., Chhabra, d., Kekic, m., Prst, i. V., Tsubakihara, m., Berry, d. Do., & Nosworthy, n. J. (2003). Vezavni proteini aktin: regulacija citoskeletnih mikrofilamentov. Fiziološki pregledi, 83(2), 433–473.
- Guo, h., Fauci, l., Shelley, m., & Kanso in. (2018). Bistabilnost pri sinhronizaciji aktiviranih mikrofilamentov. Journal of Fluid Mechanics, 836, 304-323.
- Kosilo., Langer, r., & Vaicanti, J. Str. (Eds.). (2011). Načela tkivnega inženiringa. Akademski tisk.
- Robbins, j. (2017). Bolezni citoskeleta: Deminopatije. Pri kardioskeletnih miopatijah pri otrocih in odraslih (str. 173-192). Akademski tisk.
- « Značilnosti medveda grizli, habitat, hrana, vedenje
- Značilnosti cistus clusii, taksonomija, habitat, distribucija, uporaba »