

Cinkovi prsti, struktura, klasifikacija, funkcija
- 4808
- 527
- Miguel Gutmann DVM
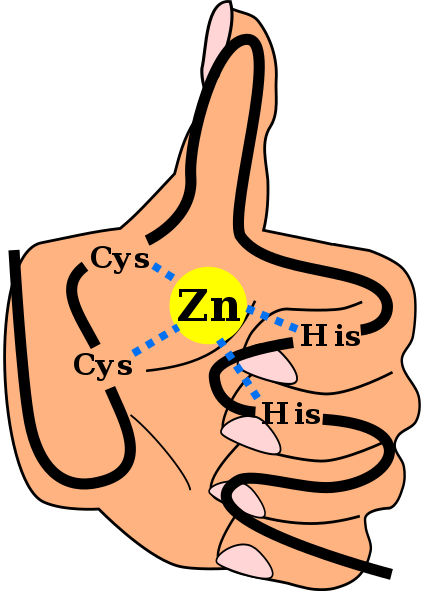
Primarna struktura domene cinkovega prsta C2H2, vključno s povezavami, ki usklajujejo cinkov ion, in z ozadjem "roke in prsta". Angel Herraez, Wikimedia Commons The Cinkovi prsti (ZF) so strukturni motivi, prisotni v velikem številu evkariotov. Spadajo v skupino metaloproteinov, saj so sposobni združiti cinkov kovinski ion, ki ga potrebujejo za njegovo delovanje. Predvideva se, da več kot 1500 domen ZF obstaja pri približno 1000 različnih beljakovin pri ljudeh.
Izraz cinkov prst ali "cinkov prst" so leta 1985 prvič skovali Miller, McLachlan in Klug, medtem ko so podrobno preučevali majhne domene DNK DNK transkripcijskega faktorja tfiiia Xenopus laevis, opisali drugi avtorji pred nekaj leti prej.
Beljakovine z motivi ZF so najpogostejše v genomu evkariontskih organizmov in sodelujejo pri raznolikosti esencialnih celičnih procesov, med katerimi izstopajo genetsko transkripcijo, prevajanje beljakovin, presnovo, zloženost in sestavljanje drugih beljakovin in lipidov, med tem pa načrtujejo celično smrt, med nji drugi.
Struktura
Struktura zf motivov je izjemno ohranjena. Običajno imajo te ponavljajoče se regije 30 do 60 aminokislin, katerih sekundarna struktura je kot dve beta antiparalni listi, ki tvorita vilice in alfa proxy, ki je označena kot ββα.
Ta sekundarna struktura se stabilizira s hidrofobnimi interakcijami in koordinacijo cinka atoma, ki jo dajeta dva cistein in dva odpadka histidina (CYS2Njegov2). Vendar obstajajo ZF, ki lahko usklajuje več kot en cinkov atom in druge, kjer se vrstni red CYS in njegovi odpadki razlikuje.
Vam lahko služi: palmitholeinska kislina: struktura, funkcije, kje jeZF -ji lahko ponovite v seriji, linearno konfigurirani v istem beljakovinah. Vsi imajo podobne strukture, vendar se lahko kemično razlikujejo z različicami ključnih aminokislinskih odpadkov za izpolnitev njihovih funkcij.
Skupna značilnost med ZF je njegova sposobnost prepoznavanja molekul DNK ali RNA različnih dolžin, zato so bili sprva šteli le za transkriptne dejavnike.
Na splošno je prepoznavanje 3PB območja v DNK in se doseže, ko protein z domeno ZF predstavlja alfa propeler do glavnega utora molekule DNK.
Klasifikacija
Obstajajo različni razlogi ZF, ki se med seboj razlikujejo glede na svojo naravo in različne vesoljske konfiguracije, ki dosegajo koordinacijske povezave z cinkovim atomom. Ena od klasifikacij je naslednja:
C2H2
To je razlog, ki ga običajno najdemo v zf. Večina razlogov c2H2 Specifični so za interakcijo z DNK in RNA, vendar so opazili sodelovanje v interakcijah beljakovin in beljakovin. Imajo med 25 in 30 aminokislinskih odpadkov in so v največji družini regulativnih beljakovin pri sesalcih.
C2H
Interakcijo z RNA in nekaterimi drugimi beljakovinami. Opazujejo jih predvsem kot del nekaterih beljakovin retrovirusnega kapsida, ki sodelujejo pri embalaži virusne RNA takoj po podvajanju.
C4 (zanka ali trak)
Beljakovine s tem motivom so encimi, ki so odgovorni za podvajanje in prepisovanje DNK. Dober primer teh so lahko surovi encimi fagov T4 in T7.
C4 (Družina Gata)
Ta družina ZF vključuje transkripcijske faktorje, ki uravnavajo izražanje pomembnih genov v številnih tkivih med razvojem celic. Dejavniki GATA-2 in 3 so na primer vključeni v hematopoezo.
Lahko vam služi: ATP (adenozin trifosfat)C6
Te domene so značilne za kvasovke, zlasti protein GAL4, ki aktivira transkripcijo genov, ki sodelujejo pri uporabi galaktoze in melibije.
Cinkovi prsti (c3Hc4-C3H2C3)
Te posebne strukture imajo 2 podtipa domene zf (c3Hc4 in c3H2C3) in so prisotni v številnih živalskih in rastlinskih beljakovinah.
Najdemo jih v beljakovinah, kot je RAD5, ki sodelujejo pri popravilu DNK v evkariontskih organizmih. Najdemo tudi v RAG1, nepogrešljivi za ponovno konfiguracijo imunoglobulinov.
H2C2
Ta domena ZF je zelo ohranjena v integralih retrovirusa in retrotransposona; Ko se pridruži belemu beljakovinam, povzroči konformacijsko spremembo v istem.
Funkcije
ZF domene beljakovine služijo različnim namenom: najdemo jih v ribosomalnih beljakovinah ali prepisni adapterji. Odkrili so tudi kot sestavni del strukture RNA polimeraze II kvasa.
Zdi se, da sodelujejo v medcelični cinkovi homeostazi in regulaciji apoptoze ali programirane celične smrti. Poleg tega obstajajo nekateri ZF beljakovine, ki delujejo kot kaperoni za zlaganje ali transport drugih beljakovin.
Lipidna zveza in temeljna vloga v interakcijah beljakovin in beljakovin so tudi izjemne funkcije domen ZF v nekaterih beljakovinah.
Biotehnološki pomen
Z leti je strukturno in funkcionalno razumevanje domen ZF omogočilo velik znanstveni napredek, ki pomeni uporabo njihovih značilnosti za biotehnološke namene.
Ker imajo nekateri ZF proteini veliko specifičnost zaradi nekaterih domen DNK, je trenutno veliko truda vloženih v specifično zasnovo ZF, ki lahko zagotavlja dragocen napredek pri genski terapiji pri ljudeh.
Lahko vam služi: Kitajska flora in favna: reprezentativna vrstaZanimive biotehnološke aplikacije izhajajo tudi iz zasnove beljakovin z ZF, ki ga spreminja genetski inženiring. Odvisno od želenega konca je mogoče nekatere spremeniti z dodajanjem prstov s prsti "poli cinkove", ki so sposobni praktično prepoznati katero koli zaporedje DNK z veliko afiniteto in specifičnostjo.
Genomska izdaja s spremenjenimi nukleaji je ena najbolj obetavnih aplikacij danes. Ta vrsta izdaje ponuja možnost izvedbe študij o genetski funkciji neposredno v zanimivem sistemu.
Genetski inženiring z uporabo spremenjenih zf nukleasa je pritegnil pozornost znanstvenikov na področju genetskega izboljšanja kultivarjev agronomskega pomena. Ta jedra so bila uporabljena za popravljanje endogenega gena, ki proizvaja herbicidne -odmevne oblike v tobačnih rastlinah.
Nucleas z ZF so bila uporabljena tudi za dodajanje genov v celicah sesalcev. Zadevni beljakovine so bile uporabljene za ustvarjanje nabora izogenih mišjih celic z nizom alelov, definiranih za endogeni gen.
Tak postopek ima neposredno uporabo pri označevanju in ustvarjanju novih alelnih oblik za preučevanje strukture in funkcijskih razmerij v pogojih iz naravnega izražanja in v izogenem okolju.
Reference
- Berg, j. M. (1990). Domene cinkovega prsta: hipoteze in trenutno znanje. Letni pregled biofizike in biofizikalne kemije, 19(39), 405–421.
- Kluska, k., Adamczyk, j., & Krȩzel, a. (2017). Kovinske vezavne lastnosti cinkovih prstov z naravnim alten kovinskim vezanjem. Metalomika, 10(2), 248-263.